

Dissociation among in vitro telomerase activity, telomere maintenance, and cellular immortalization
- PMID: 9843956
- PMCID: PMC24516
- DOI: 10.1073/pnas.95.25.14723
Dissociation among in vitro telomerase activity, telomere maintenance, and cellular immortalization
Abstract
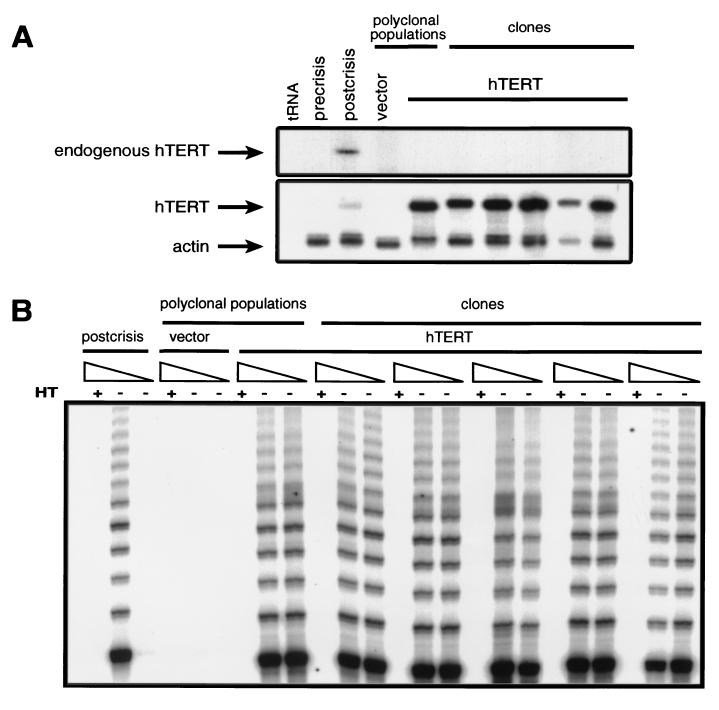
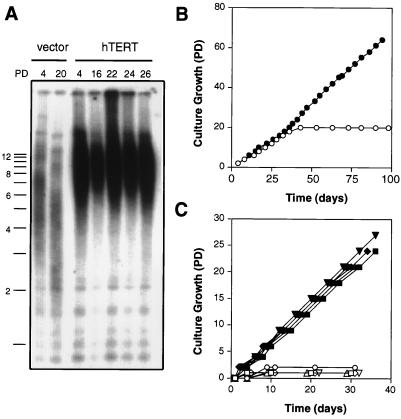
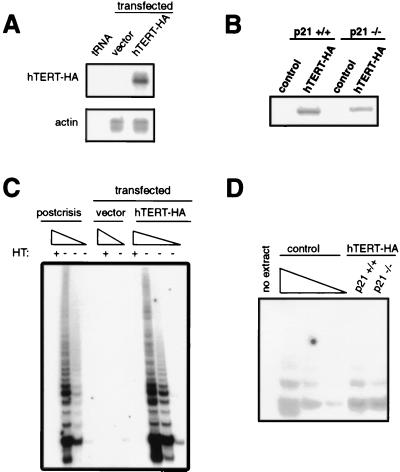
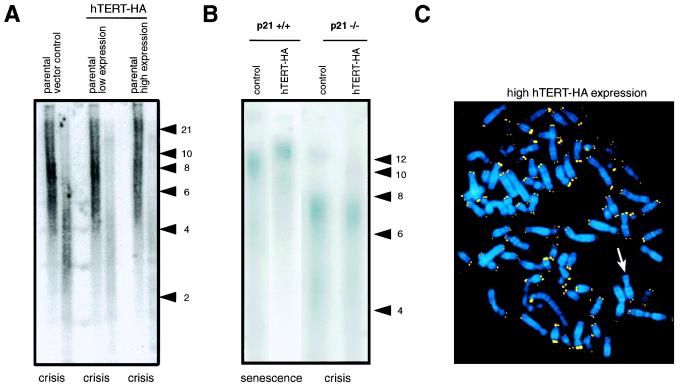
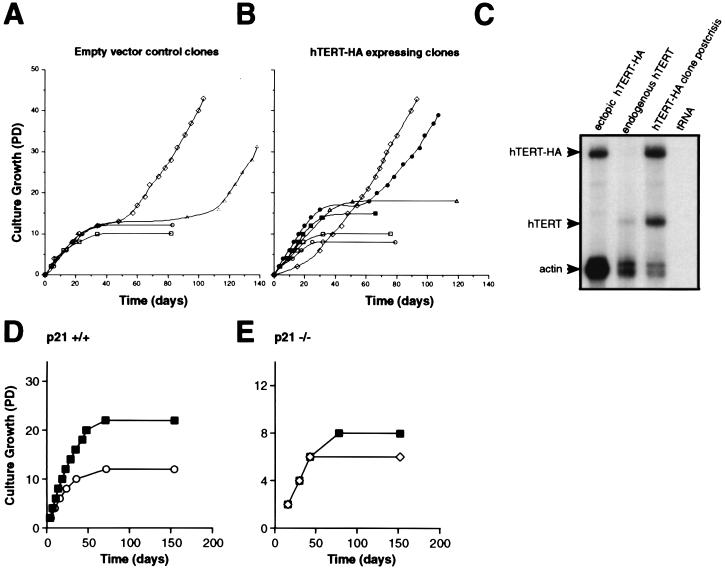
The immortalization of human cells is a critical step during tumorigenesis. In vitro, normal human somatic cells must overcome two proliferative blockades, senescence and crisis, to become immortal. Transformation with viral oncogenes extends the life span of human cells beyond senescence. Such transformed cells eventually succumb to crisis, a period of widespread cellular death that has been proposed to be the result of telomeric shortening. We now show that ectopic expression of the telomerase catalytic subunit (human telomerase reverse transcriptase or hTERT) and subsequent activation of telomerase can allow postsenescent cells to proliferate beyond crisis, the last known proliferative blockade to cellular immortality. Moreover, we demonstrate that alteration of the carboxyl terminus of human telomerase reverse transcriptase does not affect telomerase enzymatic activity but impedes the ability of this enzyme to maintain telomeres. Telomerase-positive cells expressing this mutant enzyme fail to undergo immortalization, further tightening the connection between telomere maintenance and immortalization.
Figures
Comment in
-
Crisis intervention: the role of telomerase.Proc Natl Acad Sci U S A. 1999 Mar 30;96(7):3339-41. doi: 10.1073/pnas.96.7.3339. Proc Natl Acad Sci U S A. 1999. PMID: 10097039 Free PMC article. Review. No abstract available.
References
-
- Hayflick L, Moorhead P S. Exp Cell Res. 1961;25:585–621. - PubMed
-
- Kilian A, Bowtell D D L, Abud H E, Hime G R, Venter D J, Keese P K, Duncan E L, Reddel R R, Jefferson R A. Hum Mol Genet. 1997;6:2011–2019. - PubMed
-
- Meyerson M, Counter C M, Ng Eaton E, Ellisen L W, Steiner P, Caddle S D, Ziaugra L, Beijersbergen R L, Davidoff M J, Liu Q, et al. Cell. 1997;90:785–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
