

The increased level of beta1,4-galactosyltransferase required for lactose biosynthesis is achieved in part by translational control
- PMID: 9843970
- PMCID: PMC24530
- DOI: 10.1073/pnas.95.25.14805
The increased level of beta1,4-galactosyltransferase required for lactose biosynthesis is achieved in part by translational control
Abstract
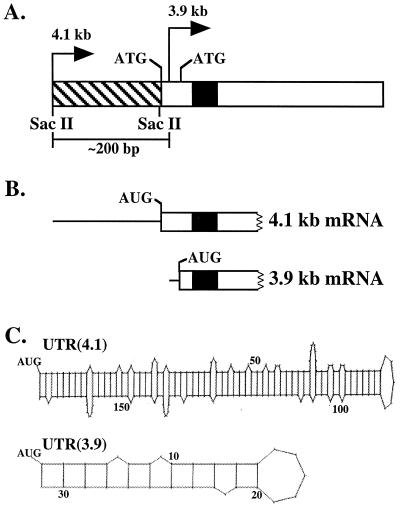
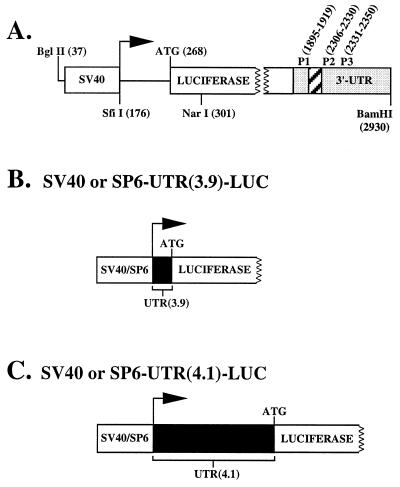
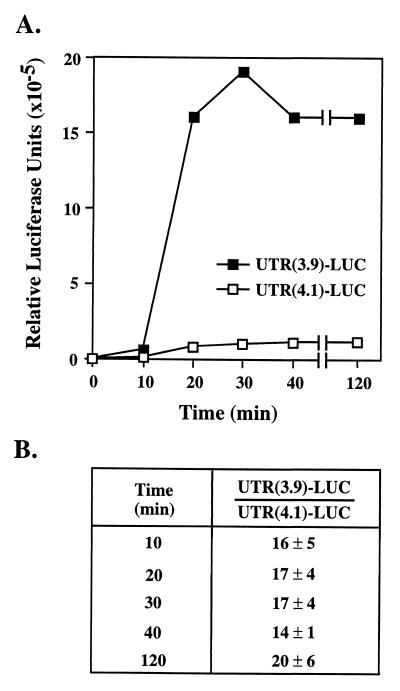
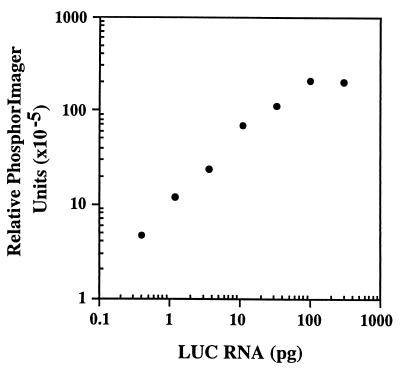
beta1,4-Galactosyltransferase (beta4GalT-I) participates in both glycoconjugate biosynthesis (ubiquitous activity) and lactose biosynthesis (mammary gland-specific activity). In somatic tissues, transcription of the mammalian beta4GalT-I gene results in a 4.1-kb mRNA and a 3.9-kb mRNA as a consequence of initiation at two start sites separated by approximately 200 bp. In the mammary gland, coincident with the increased beta4GalT-I enzyme level ( approximately 50-fold) required for lactose biosynthesis, there is a switch from the 4.1-kb start site to the preferential use of the 3.9-kb start site, which is governed by a stronger tissue-restricted promoter. The use of the 3.9-kb start site results in a beta4GalT-I transcript in which the 5'- untranslated region (UTR) has been truncated from approximately 175 nt to approximately 28 nt. The 5'-UTR of the 4.1-kb transcript [UTR(4.1)] is predicted to contain extensive secondary structure, a feature previously shown to reduce translational efficiency of an mRNA. In contrast, the 5'-UTR of the 3.9-kb mRNA [UTR(3.9)] lacks extensive secondary structure; thus, this transcript is predicted to be more efficiently translated relative to the 4.1-kb mRNA. To test this prediction, constructs were assembled in which the respective 5'-UTRs were fused to the luciferase-coding sequence and enzyme levels were determined after translation in vitro and in vivo. The luciferase mRNA containing the truncated UTR(3.9) was translated more efficiently both in vitro (approximately 14-fold) and in vivo (3- to 5-fold) relative to the luciferase mRNA containing the UTR(4.1). Consequently, in addition to control at the transcriptional level, beta4GalT-I enzyme levels are further augmented in the lactating mammary gland as a result of translational control.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
