

Models of immune memory: on the role of cross-reactive stimulation, competition, and homeostasis in maintaining immune memory
- PMID: 9843992
- PMCID: PMC24552
- DOI: 10.1073/pnas.95.25.14926
Models of immune memory: on the role of cross-reactive stimulation, competition, and homeostasis in maintaining immune memory
Abstract
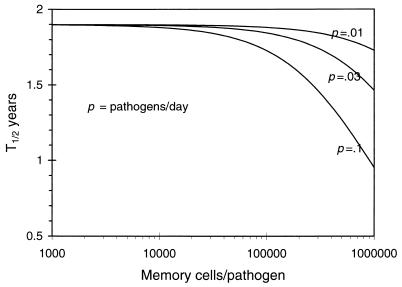
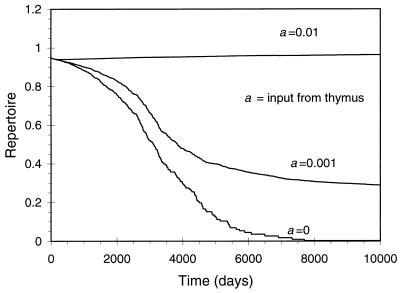
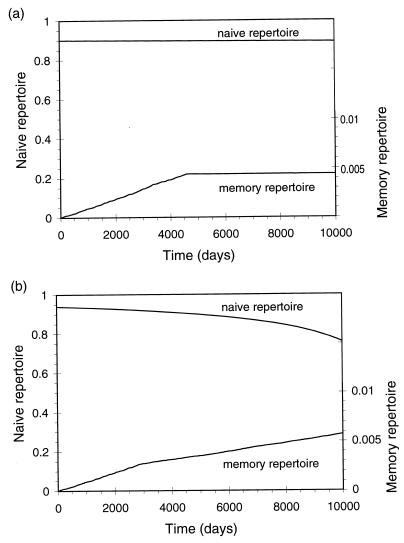
There has been much debate on the contribution of processes such as the persistence of antigens, cross-reactive stimulation, homeostasis, competition between different lineages of lymphocytes, and the rate of cell turnover on the duration of immune memory and the maintenance of the immune repertoire. We use simple mathematical models to investigate the contributions of these various processes to the longevity of immune memory (defined as the rate of decline of the population of antigen-specific memory cells). The models we develop incorporate a large repertoire of immune cells, each lineage having distinct antigenic specificities, and describe the dynamics of the individual lineages and total population of cells. Our results suggest that, if homeostatic control regulates the total population of memory cells, then, for a wide range of parameters, immune memory will be long-lived in the absence of persistent antigen (T1/2 > 1 year). We also show that the longevity of memory in this situation will be insensitive to the relative rates of cross-reactive stimulation, the rate of turnover of immune cells, and the functional form of the term for the maintenance of homeostasis.
Figures


References
-
- Hood L, Weissman I L, Wood W B, Wilson J H. Immunology. Menlo Park, California: Benjamin/Cummings; 1984.
-
- Freitas A A, Rocha B B. Immunol Today. 1993;14:25–29. - PubMed
-
- Tough D F, Sprent J. Immunol Res. 1995;14:1–12. - PubMed
-
- Tew J G, Phipps R P, Mandel T E. Immunol Rev. 1980;53:173–201. - PubMed
-
- Gray D, Skarvall H. Nature (London) 1988;336:70–73. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
