

c-Cbl/Sli-1 regulates endocytic sorting and ubiquitination of the epidermal growth factor receptor
- PMID: 9851973
- PMCID: PMC317257
- DOI: 10.1101/gad.12.23.3663
c-Cbl/Sli-1 regulates endocytic sorting and ubiquitination of the epidermal growth factor receptor
Abstract
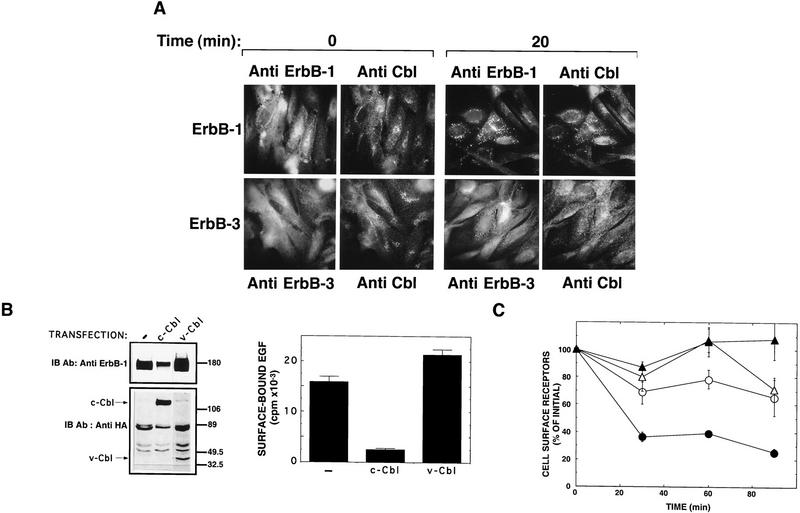
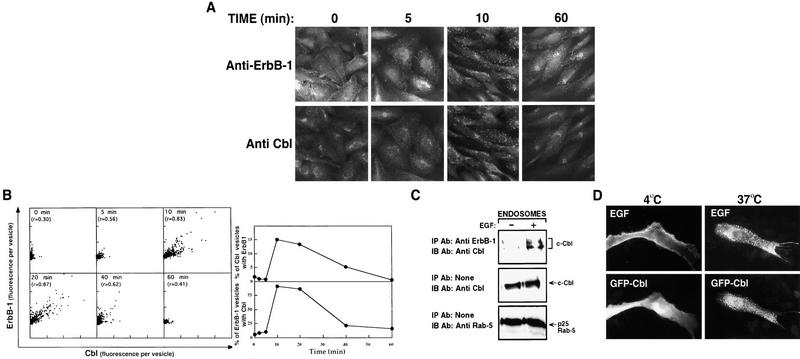
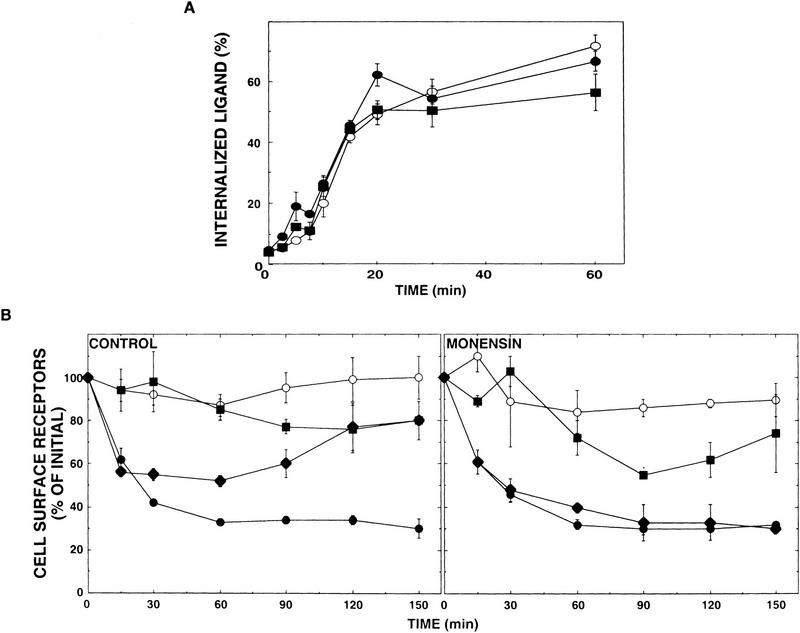
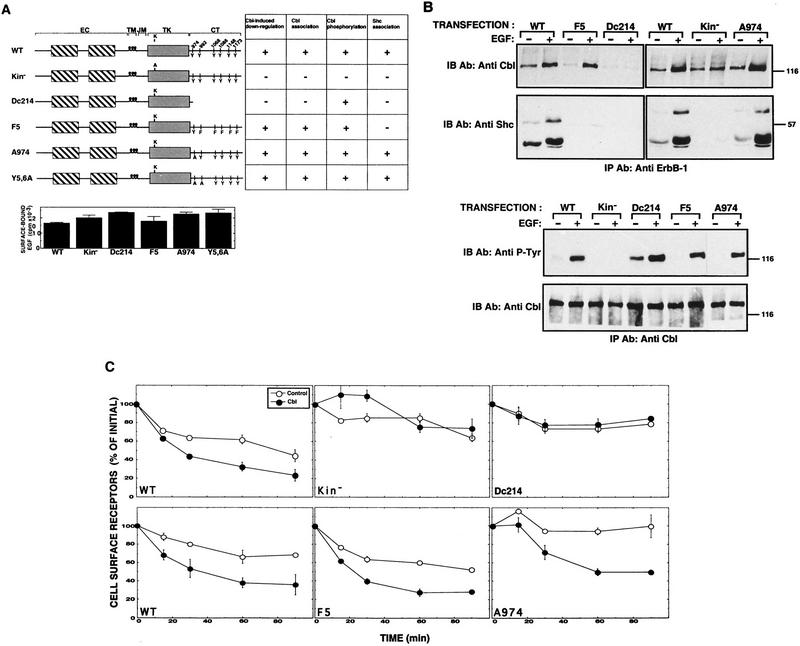
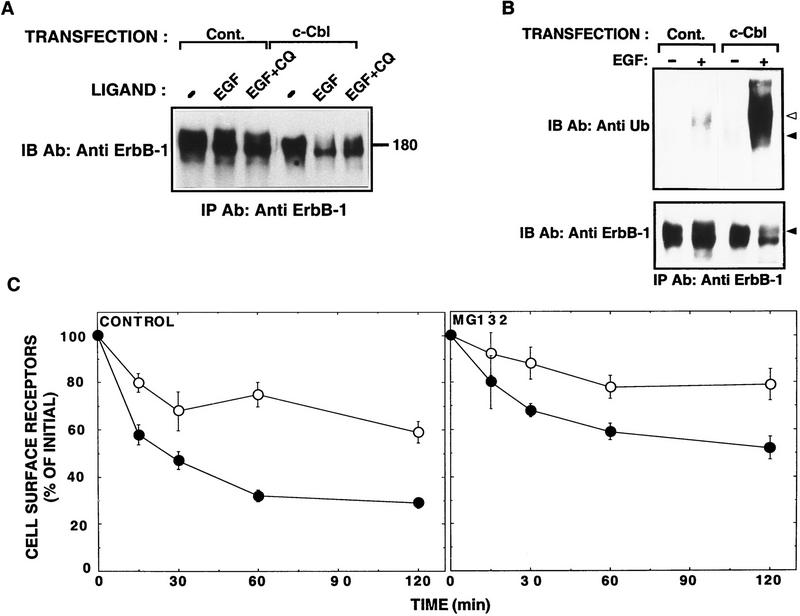
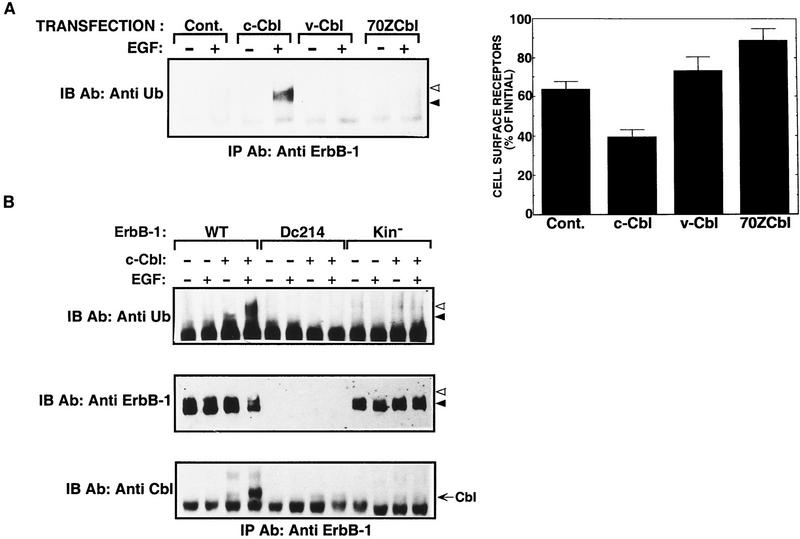
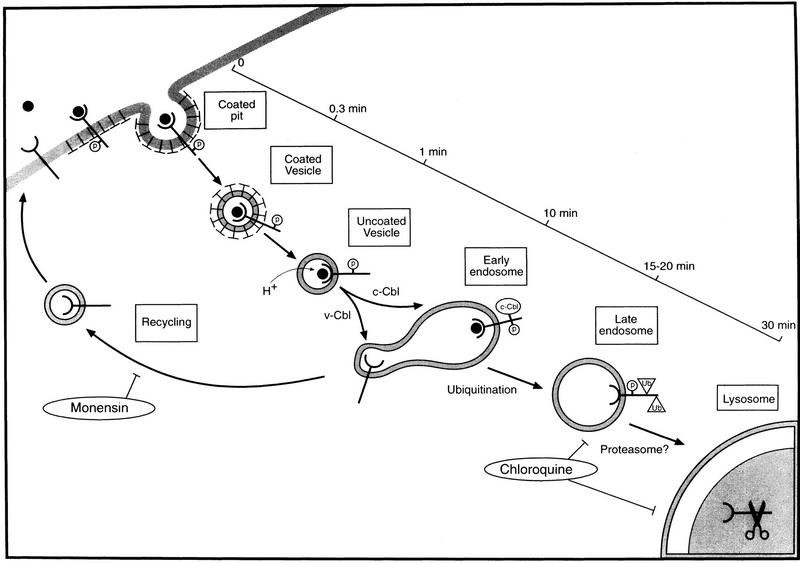
Ligand-induced down-regulation of two growth factor receptors, EGF receptor (ErbB-1) and ErbB-3, correlates with differential ability to recruit c-Cbl, whose invertebrate orthologs are negative regulators of ErbB. We report that ligand-induced degradation of internalized ErbB-1, but not ErbB-3, is mediated by transient mobilization of a minor fraction of c-Cbl into ErbB-1-containing endosomes. This recruitment depends on the receptor's tyrosine kinase activity and an intact carboxy-terminal region. The alternative fate is recycling of internalized ErbBs to the cell surface. Cbl-mediated receptor sorting involves covalent attachment of ubiquitin molecules, and subsequent lysosomal and proteasomal degradation. The oncogenic viral form of Cbl inhibits down-regulation by shunting endocytosed receptors to the recycling pathway. These results reveal an endosomal sorting machinery capable of controlling the fate, and, hence, signaling potency, of growth factor receptors.
Figures

References
-
- Alroy I, Yarden Y. The ErbB signaling network in embryogenesis and oncogenesis: Signal diversification through combinatorial ligand-receptor interactions. FEBS Lett. 1997;410:83–86. - PubMed
-
- Basu SK, Goldstein JL, Anderson RGW, Brown MS. Monensin interrupts the recycling of low density lipoprotein receptors in human fibroblasts. Cell. 1981;24:493–502. - PubMed
-
- Boussiotis VA, Freeman GJ, Berezovskaya A, Barber DL, Nadler LM. Maintenance of human T cell anergy: Blocking of IL-2 gene transcription by activated Rap1. Science. 1997;278:124–128. - PubMed
-
- Bowtell DD, Langdon WY. The protein product of the c-cbl oncogene rapidly complexes with the EGF receptor and is tyrosine phosphorylated following EGF stimulation. Oncogene. 1995;11:1561–1567. - PubMed
-
- Chang C-P, Lazar CS, Walsh BJ, Komuro M, Collawn JF, Kuhn LA, Tainer JA, Trowbridge IS, Farquhar MG, Rosenfeld MG, Wiley HS, Gill GN. Ligand-induced internalization of the epidermal growth factor receptor is mediated by multiple endocytic codes analogous to the tyrosine motif found in constitutively internalizing receptors. J Biol Chem. 1993;268:19312–19320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous