

Topological analysis of the aerobic membrane-bound formate dehydrogenase of Escherichia coli
- PMID: 9852007
- PMCID: PMC107766
- DOI: 10.1128/JB.180.24.6625-6634.1998
Topological analysis of the aerobic membrane-bound formate dehydrogenase of Escherichia coli
Abstract
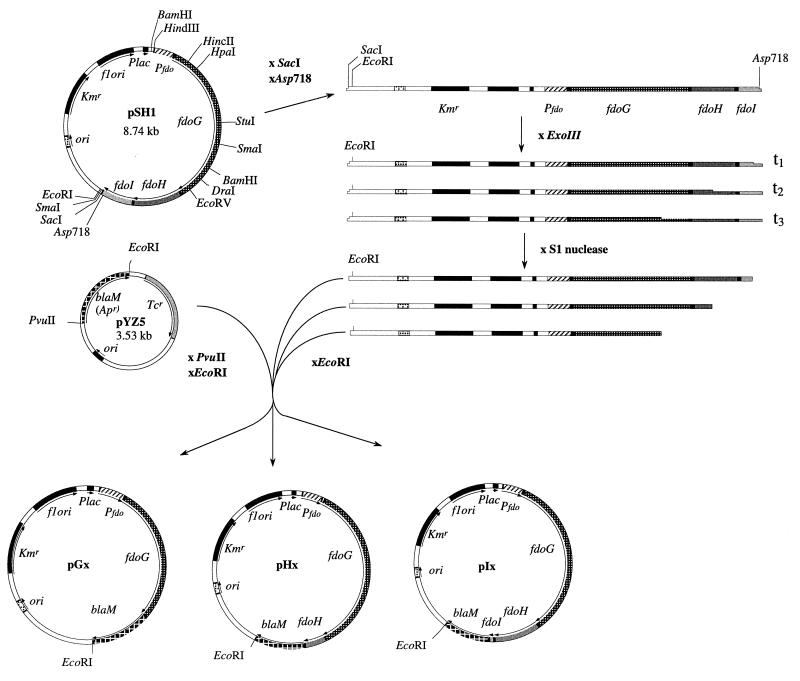
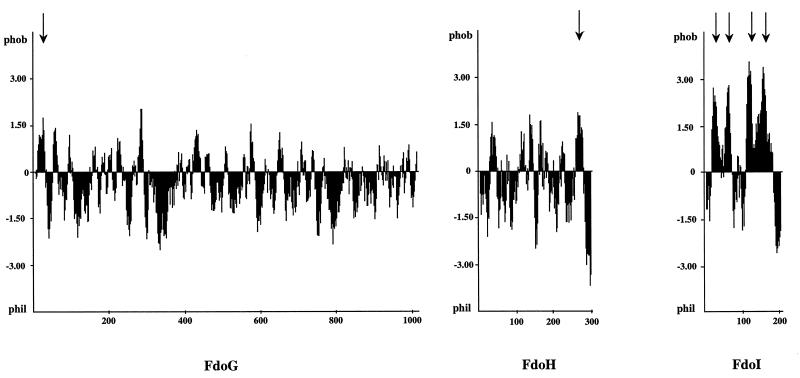
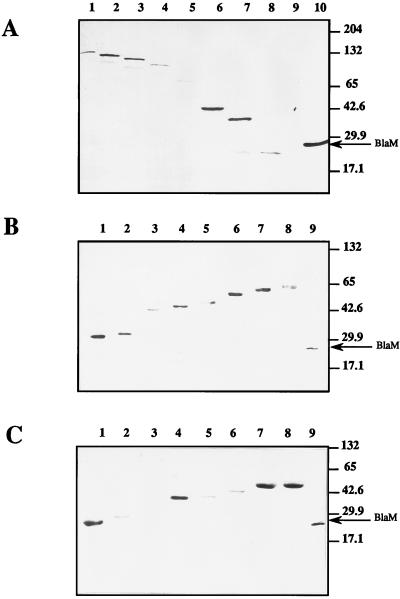
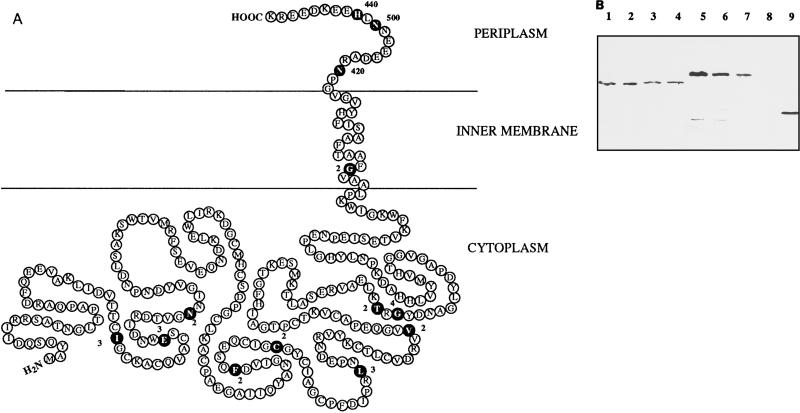
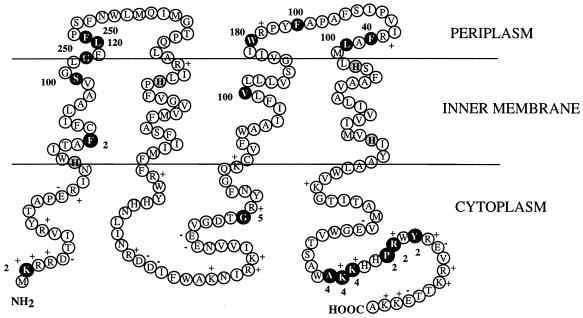
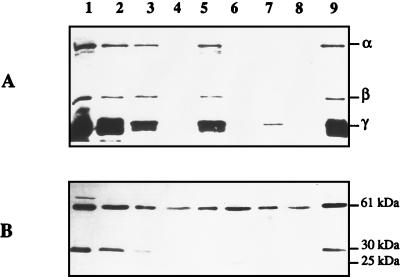
Besides formate dehydrogenase N (FDH-N), which is involved in the major anaerobic respiratory pathway in the presence of nitrate, Escherichia coli synthesizes a second isoenzyme, called FDH-O, whose physiological role is to ensure rapid adaptation during a shift from aerobiosis to anaerobiosis. FDH-O is a membrane-bound enzyme complex composed of three subunits, alpha (FdoG), beta (FdoH), and gamma (FdoI), which exhibit high sequence similarity to the equivalent polypeptides of FDH-N. The topology of these three subunits has been studied by using blaM (beta-lactamase) gene fusions. A collection of 47 different randomly generated Fdo-BlaM fusions, 4 site-specific fusions, and 3 sandwich fusions were isolated along the entire sequence of the three subunits. In contrast to previously reported predictions from sequence analysis, our data suggested that the alphabeta catalytic dimer is located in the cytoplasm, with a C-terminal anchor for beta protruding into the periplasm. As expected, the gamma subunit, which specifies cytochrome b, was shown to cross the cytoplasmic membrane four times, with the N and C termini exposed to the cytoplasm. Protease digestion studies of the 35S-labelled FDH-O heterotrimer in spheroplasts add further support to this model. Consistently, prior studies regarding the bioenergetic function of formate dehydrogenase provided evidence for a mechanism in which formate is oxidized in the cytoplasm.
Figures
References
-
- Augier V B, Guigliarelli M, Asso P, Bertrand C, Frixon G, Giordano G, Chippaux M, Blasco F. Site-directed mutagenesis of conserved cysteine residues within the β subunit of Escherichia coli nitrate reductase. Physiological, biochemical, and EPR characterization of the mutated enzymes. Biochemistry. 1993;32:2013–2023. - PubMed
-
- Ballantine S P, Boxer D H. Isolation and characterisation of a soluble active fragment of hydrogenase isoenzyme 2 from the membranes of anaerobically grown Escherichia coli. Eur J Biochem. 1986;156:277–284. - PubMed
-
- Bartolomé B, Jubete Y, Martinez E, de la Cruz F. Construction and properties of a family of pACYC184-derived cloning vectors compatible with pBR322 and its derivatives. Gene. 1991;102:75–78. - PubMed
-
- Begg Y A, Whyte J N, Haddock B A. The identification of mutants of Escherichia coli deficient in formate dehydrogenase and nitrate reductase activities using dye indicators plates. FEMS Microbiol Lett. 1977;2:47–50.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
