

Functional similarities between the Listeria monocytogenes virulence regulator PrfA and cyclic AMP receptor protein: the PrfA* (Gly145Ser) mutation increases binding affinity for target DNA
- PMID: 9852011
- PMCID: PMC107770
- DOI: 10.1128/JB.180.24.6655-6660.1998
Functional similarities between the Listeria monocytogenes virulence regulator PrfA and cyclic AMP receptor protein: the PrfA* (Gly145Ser) mutation increases binding affinity for target DNA
Abstract
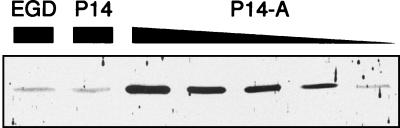
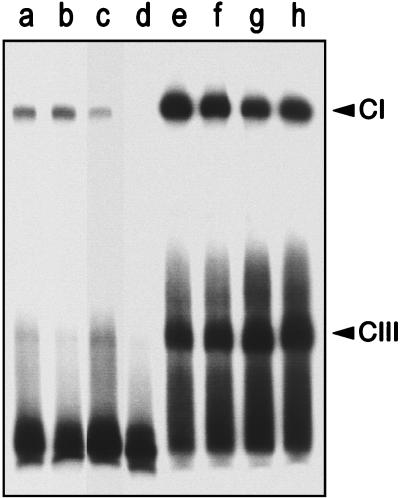
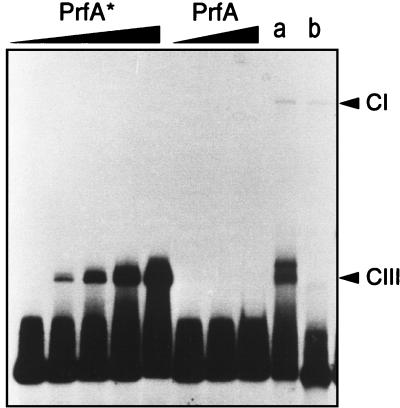
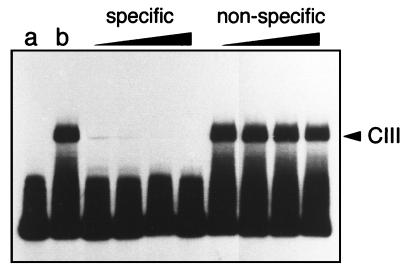
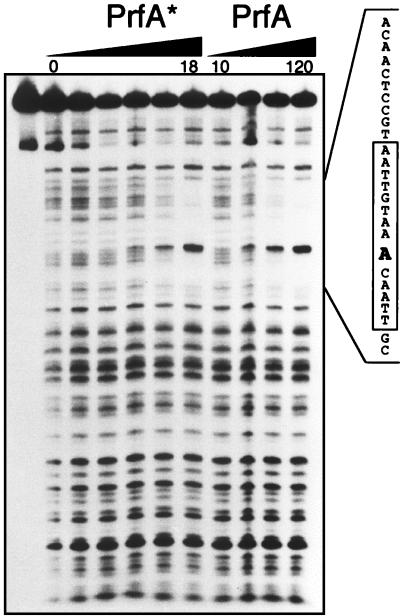
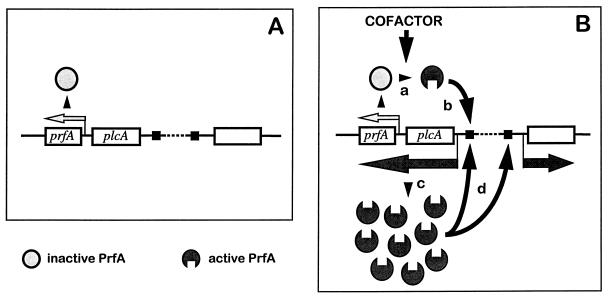
Most Listeria monocytogenes virulence genes are positively regulated by the PrfA protein, a transcription factor sharing sequence similarities with cyclic AMP (cAMP) receptor protein (CRP). Its coding gene, prfA, is regulated by PrfA itself via an autoregulatory loop mediated by the upstream PrfA-dependent plcA promoter. We have recently characterized prfA* mutants from L. monocytogenes which, as a result of a single amino acid substitution in PrfA, Gly145Ser, constitutively overexpress prfA and the genes of the PrfA virulence regulon. Here, we show that about 10 times more PrfA protein is produced in a prfA* strain than in the wild type. Thus, the phenotype of prfA* mutants is presumably due to the synthesis of a PrfA protein with higher promoter-activating activity (PrfA*), which keeps its intracellular levels constantly elevated by positive feedback. We investigated the interaction of PrfA and PrfA* (Gly145Ser) with target DNA. Gel retardation assays performed with a DNA fragment carrying the PrfA binding site of the plcA promoter demonstrated that the PrfA* mutant form is much more efficient than wild-type PrfA at forming specific DNA-protein complexes. In footprinting experiments, the two purified PrfA forms interacted with the same nucleotides at the target site, although the minimum amount required for protection was 6 to 7 times lower with PrfA*. These results show that the primary functional consequence of the Gly145Ser mutation is an increase in the affinity of PrfA for its target sequence. Interestingly, similar mutations at the equivalent position in CRP result in a transcriptionally active, CRP* mutant form which binds with high affinity to target DNA in the absence of the activating cofactor, cAMP. Our observations suggest that the structural similarities between PrfA and CRP are also functionally relevant and support a model in which the PrfA protein, like CRP, shifts from transcriptionally inactive to active conformations by interaction with a cofactor.
Figures
References
-
- Baichoo N, Heyduk T. Mapping conformational changes in a protein: application of a protein footprinting technique to cAMP-induced conformational changes in cAMP receptor protein. Biochemistry. 1997;36:10830–10836. - PubMed
-
- Blazy B, Ullmann A. Properties of cyclic AMP-independent catabolite gene activator proteins of Escherichia coli. J Biol Chem. 1986;261:11645–11649. - PubMed
-
- Böckmann R, Dickneite C, Middendorf B, Goebel W, Sokolovic Z. Specific binding of the Listeria monocytogenes transcriptional regulator PrfA to target sequences requires additional factor(s) and is influenced by iron. Mol Microbiol. 1996;22:643–653. - PubMed
-
- Bohne J, Kestler H, Uebele C, Sokolovic Z, Goebel W. Differential regulation of the virulence genes of Listeria monocytogenes by the transcriptional activator PrfA. Mol Microbiol. 1996;20:1189–1198. - PubMed
-
- Bohne J, Sokolovic Z, Goebel W. Transcriptional regulation of prfA and PrfA-regulated virulence genes in Listeria monocytogenes. Mol Microbiol. 1994;11:1141–1150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
