

Chromosomal proteins HMG-14 and HMG-17 are released from mitotic chromosomes and imported into the nucleus by active transport
- PMID: 9852141
- PMCID: PMC2132996
- DOI: 10.1083/jcb.143.6.1427
Chromosomal proteins HMG-14 and HMG-17 are released from mitotic chromosomes and imported into the nucleus by active transport
Abstract
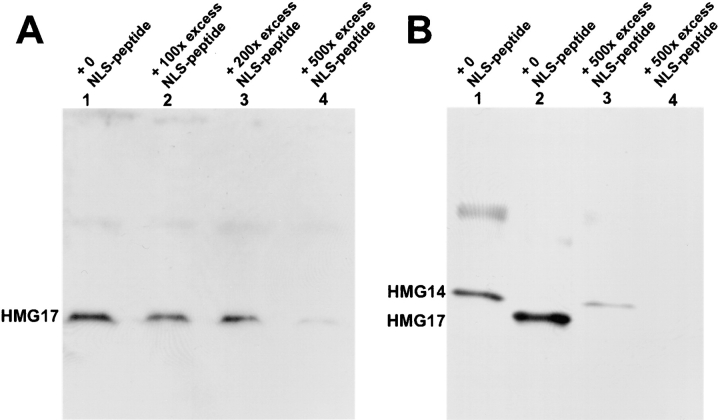
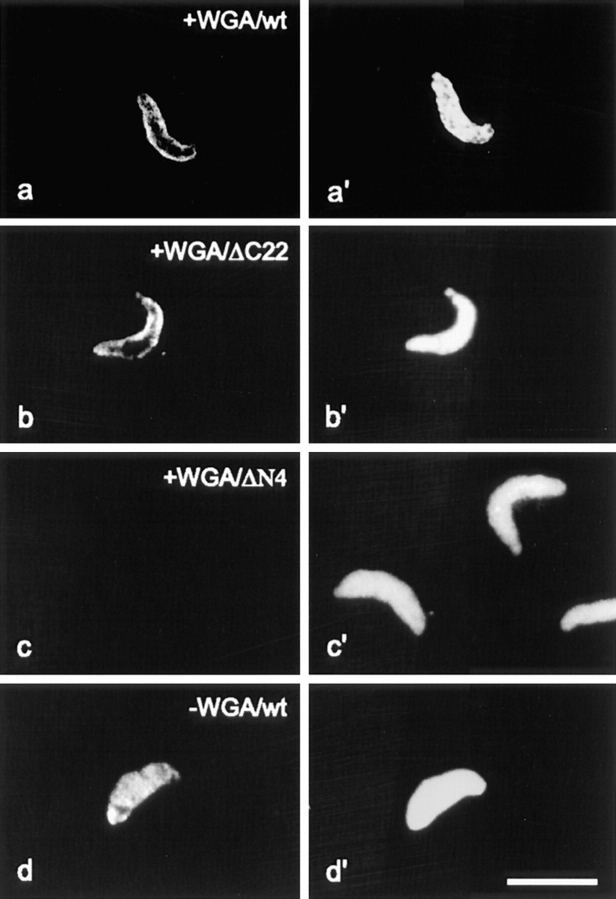
The high mobility group 14/17 (HMG-14/-17) proteins form specific complexes with nucleosome core particles and produce distinct footprints on nucleosomal DNA. Therefore, they could be an integral part of the chromatin fiber. Here we show that during the cell cycle these proteins are transiently dissociated from chromatin. They colocalize with the nuclear DNA in interphase and prophase but not in metaphase and anaphase. They relocate into the nucleus and colocalize again with the DNA in late telophase, concomitantly with the appearance of the nuclear envelope. Thus, these nucleosomal binding proteins are not always associated with chromatin. Using reconstituted nuclei and permeabilized cells, we demonstrate that these two small proteins, with a molecular mass <10 kD, are actively imported into the nucleus. We identify the major elements involved in the nuclear import of these chromosomal proteins: HMG-14/-17 proteins contain an intrinsic bipartite nuclear localization signal, and their entry into the nucleus through nuclear pores requires energy and the participation of importin alpha. These findings suggest that the cell cycle-related association of HMG-14/-17 with chromatin is dependent on, and perhaps regulated by, nuclear import processes.
Figures






References
-
- Adam S, Lobi T, Mitchell M, Gerace L. Identification of specific binding proteins for a nuclear localization sequence. Nature. 1989;337:276–279. - PubMed
-
- Alfonso PJ, Crippa MP, Hayes JJ, Bustin M. The footprint of chromosomal proteins HMG-14 and HMG-17 on chromatin subunits. J Mol Biol. 1994;236:189–198. - PubMed
-
- Bell P, Mais C, McStay B, Scheer U. Association of the nucleolar transcription factor UBF with the transcriptionally inactive rRNA genes of pronuclei and early Xenopusembryos. J Cell Sci. 1997;110:2053–2063. - PubMed
-
- Blow J, Laskey R. Initiation of replication in nuclei and purified DNA by a cell free extract of Xenopuseggs. Cell. 1986;47:577–587. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
