

Gap junction-mediated cell-cell communication modulates mouse neural crest migration
- PMID: 9852163
- PMCID: PMC2132985
- DOI: 10.1083/jcb.143.6.1725
Gap junction-mediated cell-cell communication modulates mouse neural crest migration
Abstract
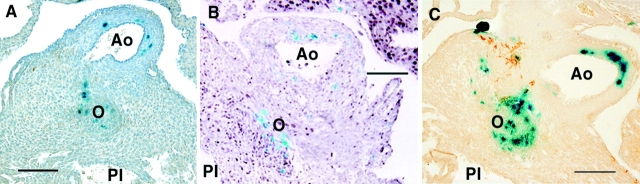
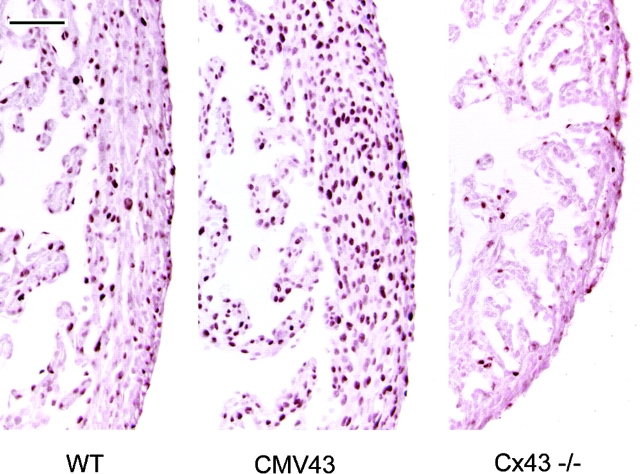
Previous studies showed that conotruncal heart malformations can arise with the increase or decrease in alpha1 connexin function in neural crest cells. To elucidate the possible basis for the quantitative requirement for alpha1 connexin gap junctions in cardiac development, a neural crest outgrowth culture system was used to examine migration of neural crest cells derived from CMV43 transgenic embryos overexpressing alpha1 connexins, and from alpha1 connexin knockout (KO) mice and FC transgenic mice expressing a dominant-negative alpha1 connexin fusion protein. These studies showed that the migration rate of cardiac neural crest was increased in the CMV43 embryos, but decreased in the FC transgenic and alpha1 connexin KO embryos. Migration changes occurred in step with connexin gene or transgene dosage in the homozygous vs. hemizygous alpha1 connexin KO and CMV43 embryos, respectively. Dye coupling analysis in neural crest cells in the outgrowth cultures and also in the living embryos showed an elevation of gap junction communication in the CMV43 transgenic mice, while a reduction was observed in the FC transgenic and alpha1 connexin KO mice. Further analysis using oleamide to downregulate gap junction communication in nontransgenic outgrowth cultures showed that this independent method of reducing gap junction communication in cardiac crest cells also resulted in a reduction in the rate of crest migration. To determine the possible relevance of these findings to neural crest migration in vivo, a lacZ transgene was used to visualize the distribution of cardiac neural crest cells in the outflow tract. These studies showed more lacZ-positive cells in the outflow septum in the CMV43 transgenic mice, while a reduction was observed in the alpha1 connexin KO mice. Surprisingly, this was accompanied by cell proliferation changes, not in the cardiac neural crest cells, but in the myocardium- an elevation in the CMV43 mice vs. a reduction in the alpha1 connexin KO mice. The latter observation suggests that cardiac neural crest cells may have a role in modulating growth and development of non-neural crest- derived tissues. Overall, these findings suggest that gap junction communication mediated by alpha1 connexins plays an important role in cardiac neural crest migration. Furthermore, they indicate that cardiac neural crest perturbation is the likely underlying cause for heart defects in mice with the gain or loss of alpha1 connexin function.
Figures
Similar articles
-
Heart and neural tube defects in transgenic mice overexpressing the Cx43 gap junction gene.Development. 1997 Apr;124(7):1281-92. doi: 10.1242/dev.124.7.1281. Development. 1997. PMID: 9118799
-
Alteration in connexin 43 gap junction gene dosage impairs conotruncal heart development.Dev Biol. 1998 Jun 1;198(1):32-44. doi: 10.1006/dbio.1998.8891. Dev Biol. 1998. PMID: 9640330
-
Heart malformations in transgenic mice exhibiting dominant negative inhibition of gap junctional communication in neural crest cells.Dev Biol. 1998 Dec 1;204(1):224-34. doi: 10.1006/dbio.1998.9089. Dev Biol. 1998. PMID: 9851855
-
Gap junction communication and the modulation of cardiac neural crest cells.Trends Cardiovasc Med. 1999 Apr-May;9(3-4):63-9. doi: 10.1016/s1050-1738(99)00015-8. Trends Cardiovasc Med. 1999. PMID: 10578519 Review.
-
Molecular determinants of neural crest migration.Am J Med Genet. 2000 Winter;97(4):280-8. doi: 10.1002/1096-8628(200024)97:4<280::aid-ajmg1278>3.0.co;2-n. Am J Med Genet. 2000. PMID: 11376439 Review.
Cited by
-
microRNAs Involved in Regulating Spontaneous Recovery in Embolic Stroke Model.PLoS One. 2013 Jun 18;8(6):e66393. doi: 10.1371/journal.pone.0066393. Print 2013. PLoS One. 2013. PMID: 23823624 Free PMC article.
-
Connexin 43 controls the multipolar phase of neuronal migration to the cerebral cortex.Proc Natl Acad Sci U S A. 2012 May 22;109(21):8280-5. doi: 10.1073/pnas.1205880109. Epub 2012 May 7. Proc Natl Acad Sci U S A. 2012. PMID: 22566616 Free PMC article.
-
Silencing or amplification of endocannabinoid signaling in blastocysts via CB1 compromises trophoblast cell migration.J Biol Chem. 2012 Sep 14;287(38):32288-97. doi: 10.1074/jbc.M112.381145. Epub 2012 Jul 24. J Biol Chem. 2012. PMID: 22833670 Free PMC article.
-
Comparison of connexin expression patterns in the developing mouse heart and human foetal heart.Mol Cell Biochem. 2003 Jan;242(1-2):121-7. Mol Cell Biochem. 2003. PMID: 12619874
-
Cx43-Associated Secretome and Interactome Reveal Synergistic Mechanisms for Glioma Migration and MMP3 Activation.Front Neurosci. 2019 Mar 19;13:143. doi: 10.3389/fnins.2019.00143. eCollection 2019. Front Neurosci. 2019. PMID: 30941001 Free PMC article.
References
-
- Bancroft M, Bellairs R. The neural crest cells of the trunk region of the chick embryo studied by SEM and TEM. ZOON (Upps) 1976;4:73–85.
-
- Bennett MVL, Barrio LC, Bargiello TA, Spray DC, Hertzberg E, Saez JC. Gap junctions: new tools, new answer, new questions. Neuron. 1991;6:305–320. - PubMed
-
- Bockman DE, Kirby ML. Dependence of thymus development on derivatives of the neural crest. Science. 1984;223:498–500. - PubMed
-
- Bruzzone R, White TW, Paul DL. Connections with connexins: the molecular basis of direct intercellular signaling. Eur J Biochem. 1996;238:1–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
