

Changes in conformation of myosin heads during the development of isometric contraction and rapid shortening in single frog muscle fibres
- PMID: 9852315
- PMCID: PMC2269081
- DOI: 10.1111/j.1469-7793.1999.305ae.x
Changes in conformation of myosin heads during the development of isometric contraction and rapid shortening in single frog muscle fibres
Abstract
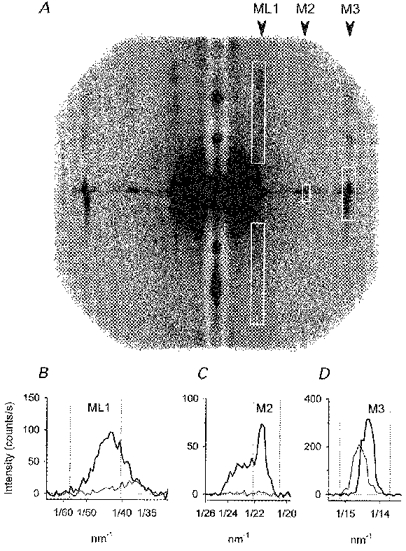
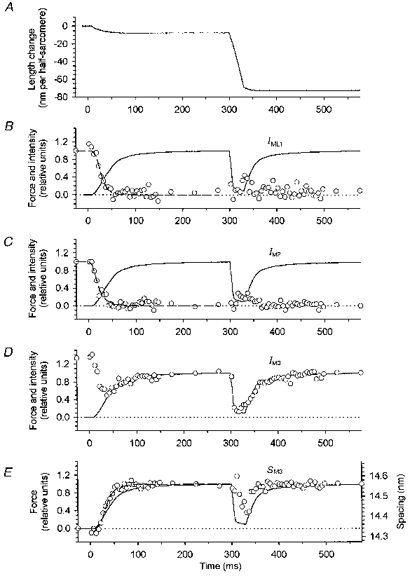
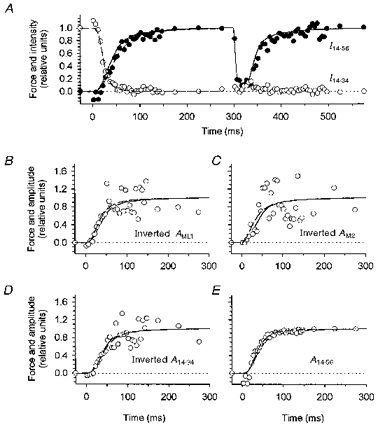
1. Two-dimensional X-ray diffraction patterns were recorded at the European Synchrotron Radiation Facility from central segments of intact single muscle fibres of Rana temporaria with 5 ms time resolution during the development of isometric contraction. Shortening at ca 0.8 times the maximum velocity was also imposed at the isometric tetanus plateau. 2. The first myosin-based layer line (ML1) and the second myosin-based meridional reflection (M2), which are both strong in resting muscle, were completely abolished at the plateau of the isometric tetanus. The third myosin-based meridional reflection (M3), arising from the axial repeat of the myosin heads along the filaments, remained intense but its spacing changed from 14.34 to 14.56 nm. The intensity change of the M3 reflection, IM3, could be explained as the sum of two components, I14.34 and I14.56, arising from myosin head conformations characteristic of rest and isometric contraction, respectively. 3. The amplitudes (A) of the X-ray reflections, which are proportional to the fraction of myosin heads in each conformation, changed with half-times that were similar to that of isometric force development, which was 33.5 +/- 2. 0 ms (mean +/- s.d., 224 tetani from three fibres, 4 C), measured from the end of the latent period. We conclude that the myosin head conformation changes synchronously with force development, at least within the 5 ms time resolution of these measurements. 4. The changes in the X-ray reflections during rapid shortening have two temporal components. The rapid decrease in intensity of the 14.56 nm reflection at the start of shortening is likely to be due to tilting of myosin heads attached to actin. The slower changes in the other reflections were consistent with a return to the resting conformation of the myosin heads that was about 60 % complete after shortening of 70 nm per half-sarcomere.
Figures
Similar articles
-
Structural changes in the myosin filament and cross-bridges during active force development in single intact frog muscle fibres: stiffness and X-ray diffraction measurements.J Physiol. 2006 Dec 15;577(Pt 3):971-84. doi: 10.1113/jphysiol.2006.115394. Epub 2006 Sep 21. J Physiol. 2006. PMID: 16990403 Free PMC article.
-
X-ray diffraction evidence for the extensibility of actin and myosin filaments during muscle contraction.Biophys J. 1994 Dec;67(6):2422-35. doi: 10.1016/S0006-3495(94)80729-5. Biophys J. 1994. PMID: 7779179 Free PMC article.
-
The structural basis of the increase in isometric force production with temperature in frog skeletal muscle.J Physiol. 2005 Sep 1;567(Pt 2):459-69. doi: 10.1113/jphysiol.2005.089672. Epub 2005 Jun 16. J Physiol. 2005. PMID: 15961426 Free PMC article.
-
X-ray diffraction studies of the contractile mechanism in single muscle fibres.Philos Trans R Soc Lond B Biol Sci. 2004 Dec 29;359(1452):1883-93. doi: 10.1098/rstb.2004.1557. Philos Trans R Soc Lond B Biol Sci. 2004. PMID: 15647164 Free PMC article. Review.
-
Two-dimensional time resolved X-ray diffraction of muscle: recent results.Adv Biophys. 1991;27:15-33. doi: 10.1016/0065-227x(91)90005-x. Adv Biophys. 1991. PMID: 1755357 Review.
Cited by
-
Structural changes in the myosin filament and cross-bridges during active force development in single intact frog muscle fibres: stiffness and X-ray diffraction measurements.J Physiol. 2006 Dec 15;577(Pt 3):971-84. doi: 10.1113/jphysiol.2006.115394. Epub 2006 Sep 21. J Physiol. 2006. PMID: 16990403 Free PMC article.
-
Impact of familial hypertrophic cardiomyopathy-linked mutations in the NH2 terminus of the RLC on β-myosin cross-bridge mechanics.J Appl Physiol (1985). 2014 Dec 15;117(12):1471-7. doi: 10.1152/japplphysiol.00798.2014. Epub 2014 Oct 16. J Appl Physiol (1985). 2014. PMID: 25324513 Free PMC article.
-
Structural changes of cross-bridges on transition from isometric to shortening state in frog skeletal muscle.Biophys J. 2006 Dec 1;91(11):4110-20. doi: 10.1529/biophysj.106.087502. Epub 2006 Sep 15. Biophys J. 2006. PMID: 16980365 Free PMC article.
-
A combined mechanical and X-ray diffraction study of stretch potentiation in single frog muscle fibres.J Physiol. 2000 Aug 1;526 Pt 3(Pt 3):589-96. doi: 10.1111/j.1469-7793.2000.00589.x. J Physiol. 2000. PMID: 10922010 Free PMC article.
-
Structural changes in myosin motors and filaments during relaxation of skeletal muscle.J Physiol. 2009 Sep 15;587(Pt 18):4509-21. doi: 10.1113/jphysiol.2009.176222. Epub 2009 Aug 3. J Physiol. 2009. PMID: 19651765 Free PMC article.
References
-
- Bordas J, Diakun GP, Diaz FG, Harries JE, Lewis RA, Lowy J, Mant GR, Martin-Fernandez ML, Towns-Andrews E. Two-dimensional time-resolved X-ray diffraction studies of live isometrically contracting frog sartorius muscle. Journal of Muscle Research and Cell Motility. 1993;14:311–324. - PubMed
-
- Burton K. Myosin step size: estimates from motility assays and shortening muscle. Journal of Muscle Research and Cell Motility. 1992;13:590–607. - PubMed
-
- Cecchi G, Griffiths PJ, Taylor S. Muscular contraction: kinetics of crossbridge attachment studied by high-frequency stiffness measurements. Science. 1982;217:70–72. - PubMed
-
- Dobbie I, Linari M, Piazzesi G, Reconditi M, Koubassova N, Ferenczi MA, Lombardi V, Irving M. Elastic bending and active tilting of myosin heads during muscle contraction. Nature. 1998;396:383–387. 10.1038/24647. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources