

Evidence that multiple P2X purinoceptors are functionally expressed in rat supraoptic neurones
- PMID: 9852319
- PMCID: PMC2269088
- DOI: 10.1111/j.1469-7793.1999.351ae.x
Evidence that multiple P2X purinoceptors are functionally expressed in rat supraoptic neurones
Abstract
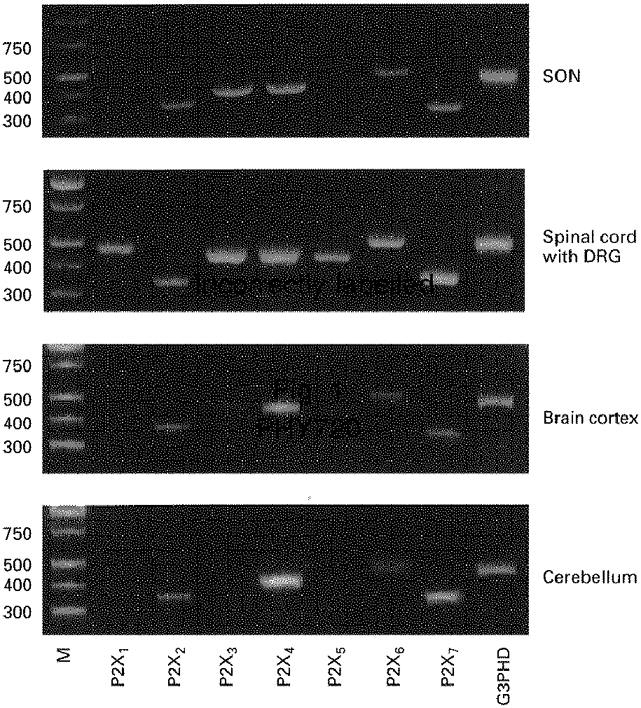
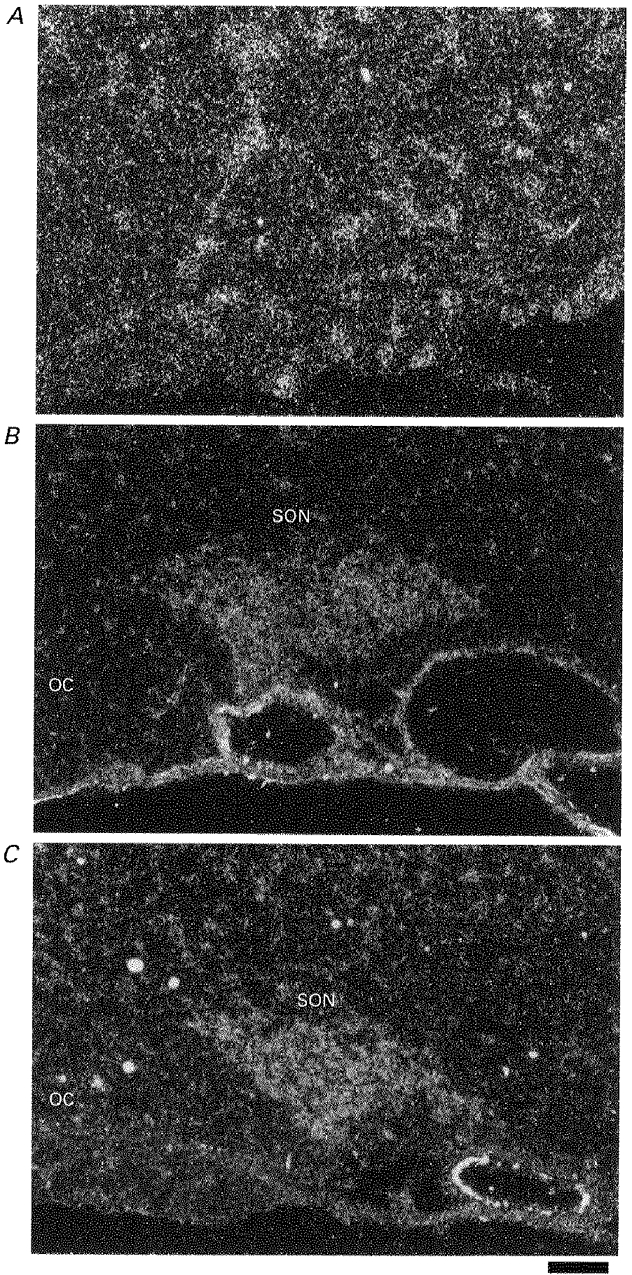
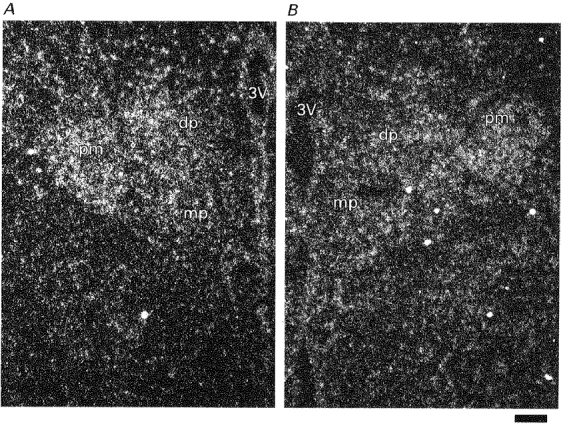
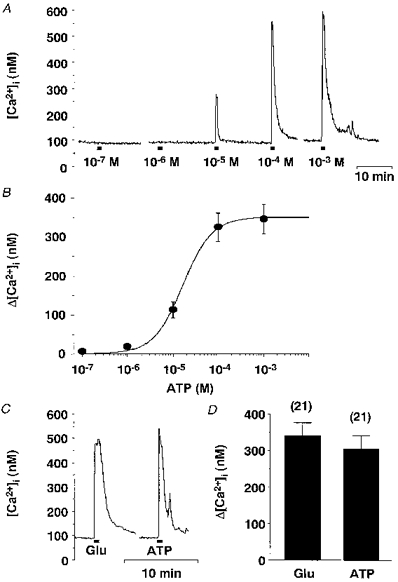
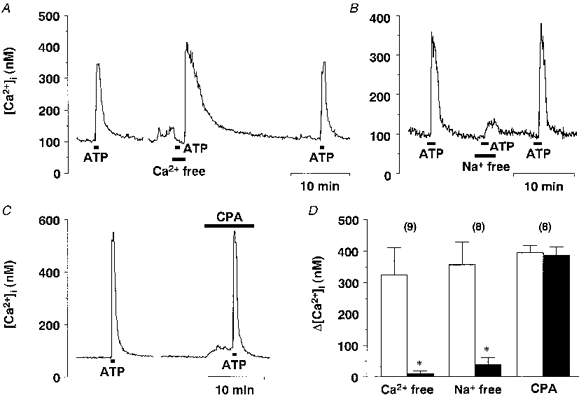
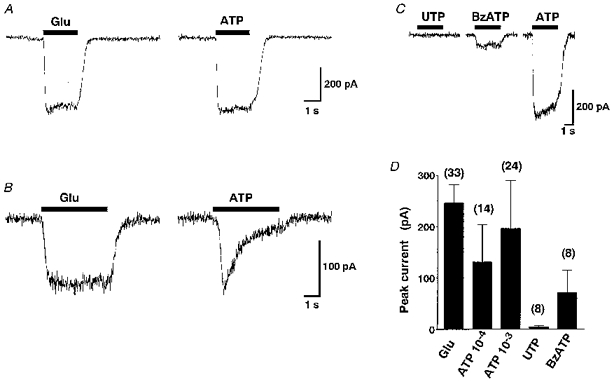
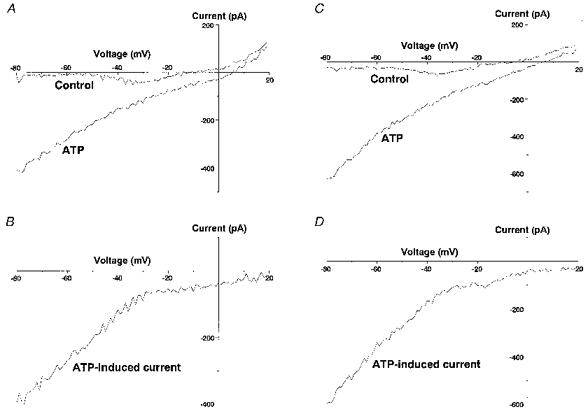
1. The expression, distribution and function of P2X purinoceptors in the supraoptic nucleus (SON) were investigated by reverse transcription-polymerase chain reaction (RT-PCR), in situ hybridization, and Ca2+-imaging and whole-cell patch-clamp techniques, respectively. 2. RT-PCR analysis of all seven known P2X receptor mRNAs in circular punches of the SON revealed that mRNAs for P2X2, P2X3, P2X4, P2X6 and P2X7 receptors were expressed in the SON, and mRNAs for P2X3, P2X4 and P2X7 were predominant. 3. In situ hybridization histochemistry for P2X3 and P2X4 receptor mRNAs showed that both mRNAs were expressed throughout the SON and in the paraventricular nucleus (PVN). 4. ATP caused an increase in [Ca2+]i in a dose-dependent manner with an ED50 of 1.7 x 10-5 M. The effects of ATP were mimicked by ATPgammaS and 2-methylthio ATP (2MeSATP), but not by AMP, adenosine, UTP or UDP. alphabeta-Methylene ATP (alphabetaMeATP) and ADP caused a small increase in [Ca2+]i in a subset of SON neurones. 5. The P2X7 agonist 2'- & 3'-O-(4-benzoylbenzoyl)-ATP (BzATP) at 10-4 M increased [Ca2+]i, but the potency of BzATP was lower than that of ATP. In contrast, BzATP caused a more prominent [Ca2+]i increase than ATP in non-neuronal cells in the SON. 6. The effects of ATP were abolished by extracellular Ca2+ removal or by the P2 antagonist pyridoxal phosphate-6-azophenyl-2',4'-disulphonic acid (PPADS), and inhibited by extracellular Na+ replacement or another P2 antagonist, suramin, but were unaffected by the P2X7 antagonist oxidized ATP, and the inhibitor of Ca2+-ATPase in intracellular Ca2+ stores cyclopiazonic acid. 7. Two patterns of desensitization were observed in the [Ca2+]i response to repeated applications of ATP: some neurones showed little or moderate desensitization, while others showed strong desensitization. 8. Whole-cell patch-clamp analysis showed that ATP induced cationic currents with marked inward rectification. The ATP-induced currents exhibited two patterns of desensitization similar to those observed in the [Ca2+]i response. 9. The results suggest that multiple P2X receptors, including P2X3, are functionally expressed in SON neurones, and that activation of these receptors induces cationic currents and Ca2+ entry. Such ionic and Ca2+-signalling mechanisms triggered by ATP may play an important role in the regulation of SON neurosecretory cells.
Figures

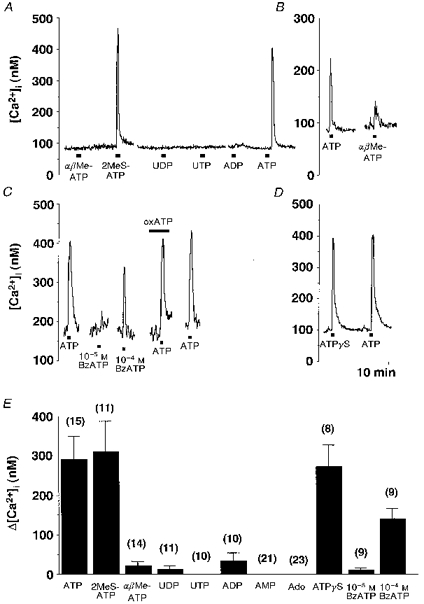
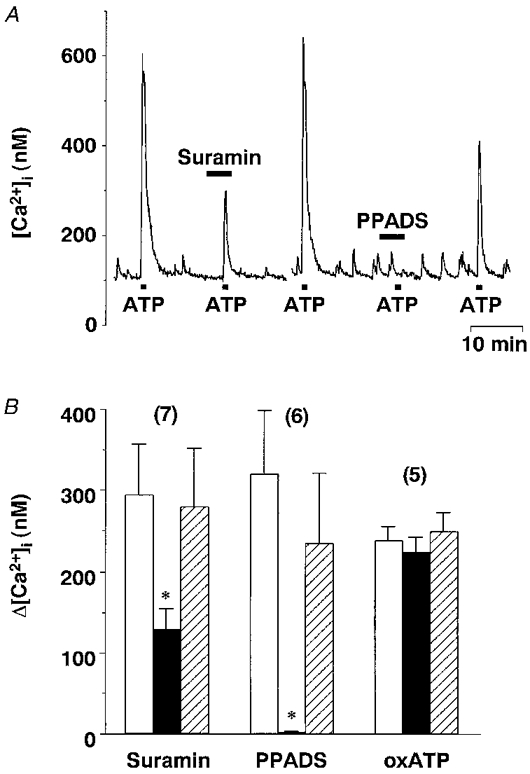
 ) and ATP plus the P2 antagonist (▪). Post-controls were obtained 10-20 min after withdrawal of each antagonist. * Significantly different (P < 0.05) from the mean of the pre- and post-control values obtained with ATP alone.
) and ATP plus the P2 antagonist (▪). Post-controls were obtained 10-20 min after withdrawal of each antagonist. * Significantly different (P < 0.05) from the mean of the pre- and post-control values obtained with ATP alone.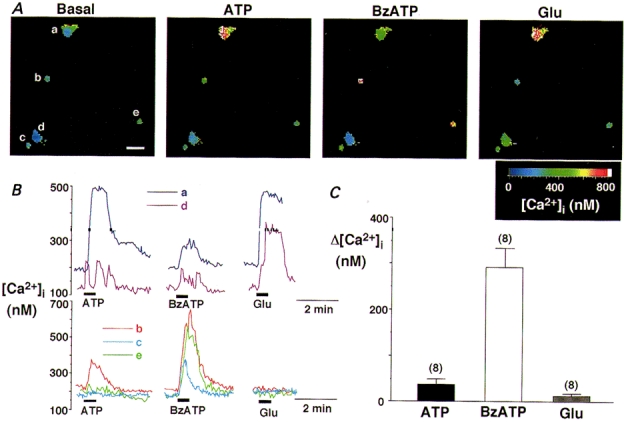
References
-
- Bo X, Zhang Y, Nassar M, Burnstock G, Schoepfer R. A P2X purinoceptor cDNA conferring a novel pharmacological profile. FEBS Letters. 1995;375:129–133. - PubMed
-
- Buller KM, Khanna S, Sibbald JR, Day TA. Central noradrenergic neurons signal via ATP to elicit vasopressin responses to haemorrhage. Neuroscience. 1996;73:637–642. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous