

Glutamate induces phosphorylation of Elk-1 and CREB, along with c-fos activation, via an extracellular signal-regulated kinase-dependent pathway in brain slices
- PMID: 9858538
- PMCID: PMC83872
- DOI: 10.1128/MCB.19.1.136
Glutamate induces phosphorylation of Elk-1 and CREB, along with c-fos activation, via an extracellular signal-regulated kinase-dependent pathway in brain slices
Abstract
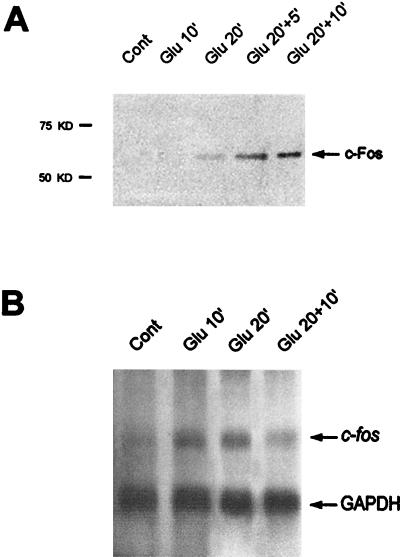
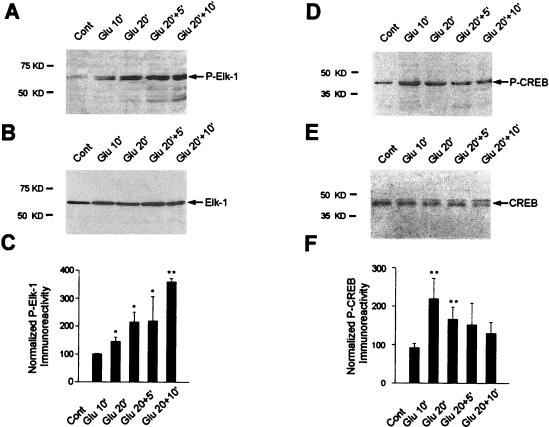
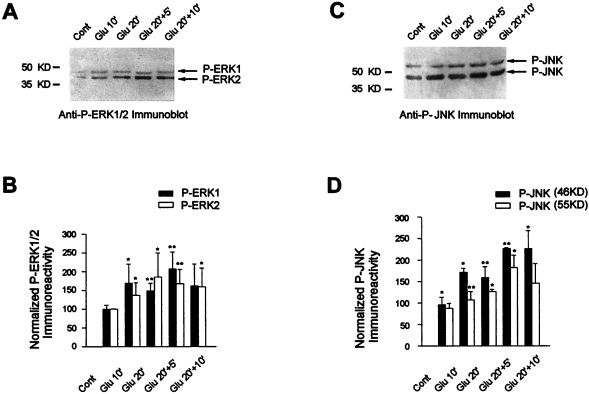
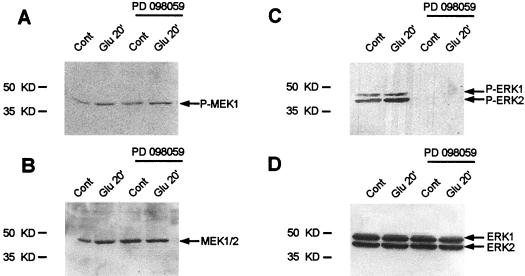
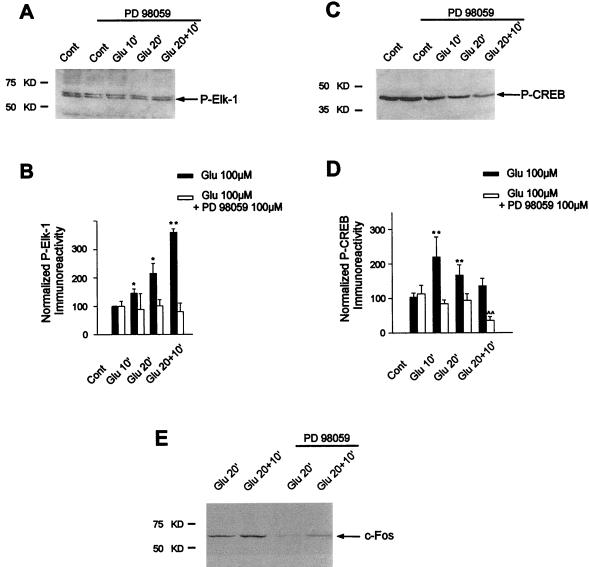
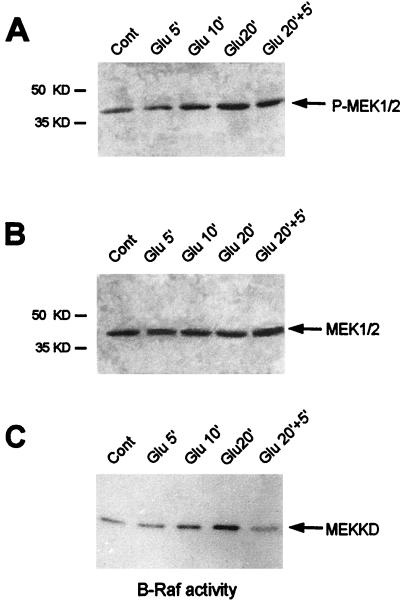
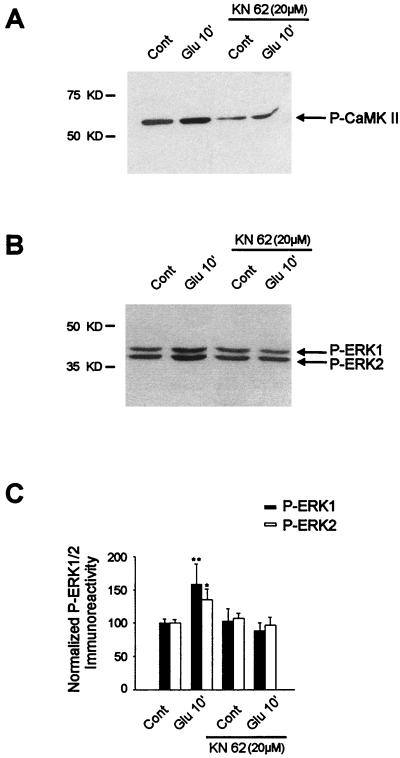
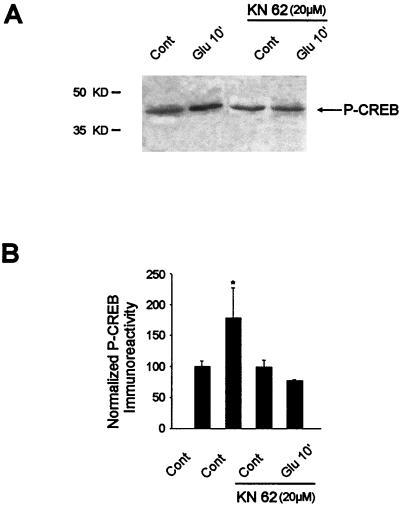
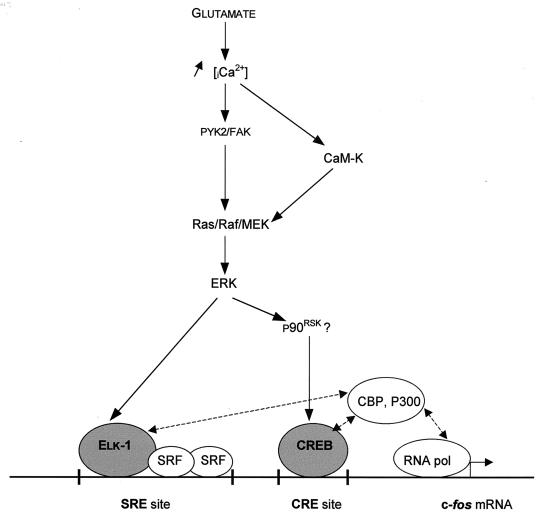
In cell culture systems, the TCF Elk-1 represents a convergence point for extracellular signal-related kinase (ERK) and c-Jun N-terminal kinase/stress-activated protein kinase (JNK/SAPK) subclasses of mitogen-activated protein kinase (MAPK) cascades. Its phosphorylation strongly potentiates its ability to activate transcription of the c-fos promoter through a ternary complex assembled on the c-fos serum response element. In rat brain postmitotic neurons, Elk-1 is strongly expressed (V. Sgambato, P. Vanhoutte, C. Pagès, M. Rogard, R. A. Hipskind, M. J. Besson, and J. Caboche, J. Neurosci. 18:214-226, 1998). However, its physiological role in these postmitotic neurons remains to be established. To investigate biochemically the signaling pathways targeting Elk-1 and c-fos in mature neurons, we used a semi-in vivo system composed of brain slices stimulated with the excitatory neurotransmitter glutamate. Glutamate treatment leads to a robust, progressive activation of the ERK and JNK/SAPK MAPK cascades. This corresponds kinetically to a significant increase in Ser383-phosphorylated Elk-1 and the appearance of c-fos mRNA. Glutamate also causes increased levels of Ser133-phosphorylated cyclic AMP-responsive element-binding protein (CREB) but only transiently relative to Elk-1 and c-fos. ERK and Elk-1 phosphorylation are blocked by the MAPK kinase inhibitor PD98059, indicating the primary role of the ERK cascade in mediating glutamate signaling to Elk-1 in the rat striatum in vivo. Glutamate-mediated CREB phosphorylation is also inhibited by PD98059 treatment. Interestingly, KN62, which interferes with calcium-calmodulin kinase (CaM-K) activity, leads to a reduction of glutamate-induced ERK activation and of CREB phosphorylation. These data indicate that ERK functions as a common component in two signaling pathways (ERK/Elk-1 and ERK/?/CREB) converging on the c-fos promoter in postmitotic neuronal cells and that CaM-Ks act as positive regulators of these pathways.
Figures
References
-
- Bading H, Ginty D D, Greenberg M E. Regulation of gene expression in hippocampal neurons by distinct signaling pathways. Science. 1993;260:181–186. - PubMed
-
- Barnier J V, Papin C, Eychène A, Lecoq O, Calothy G. The mouse B-raf gene encodes multiple protein isoforms with tissue-specific expression. J Biol Chem. 1995;270:23381–23389. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous