

Histone acetyltransferase and protein kinase activities copurify with a putative Xenopus RNA polymerase I holoenzyme self-sufficient for promoter-dependent transcription
- PMID: 9858602
- PMCID: PMC83936
- DOI: 10.1128/MCB.19.1.796
Histone acetyltransferase and protein kinase activities copurify with a putative Xenopus RNA polymerase I holoenzyme self-sufficient for promoter-dependent transcription
Abstract
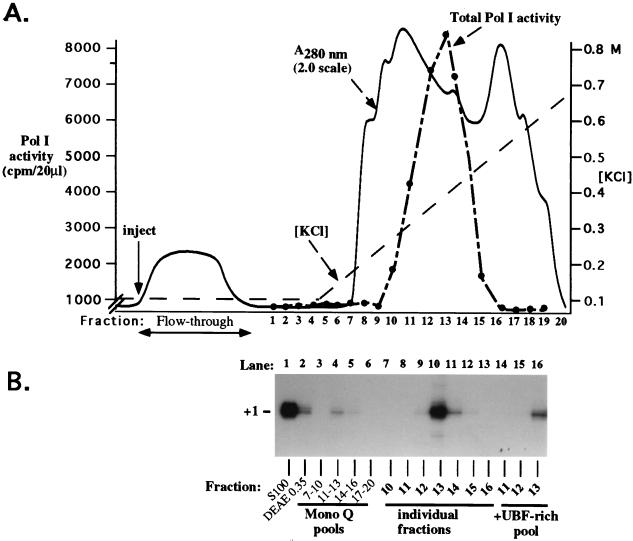
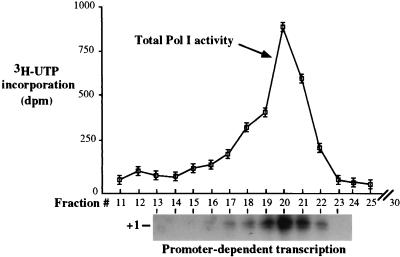
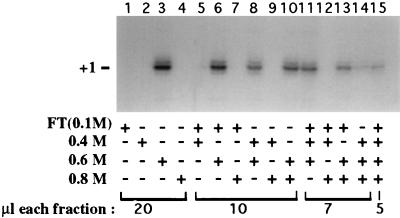
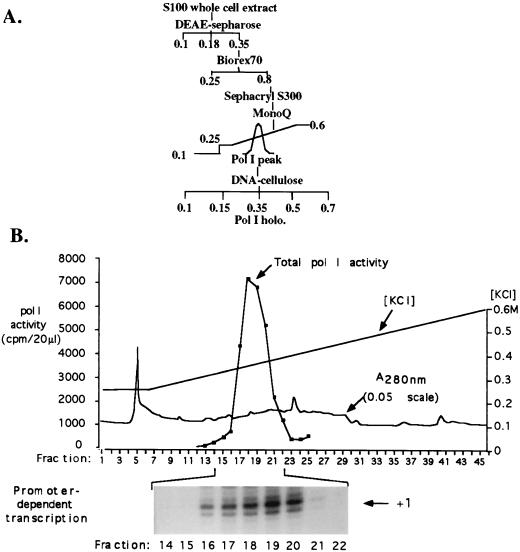
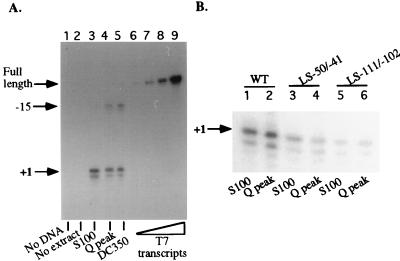
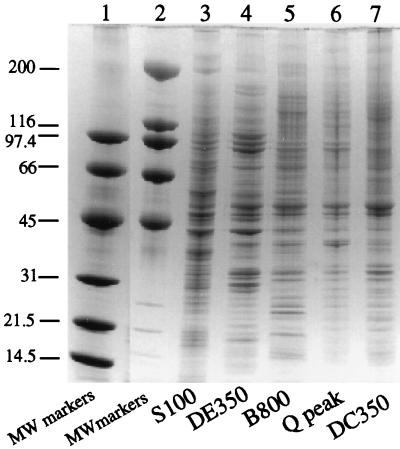
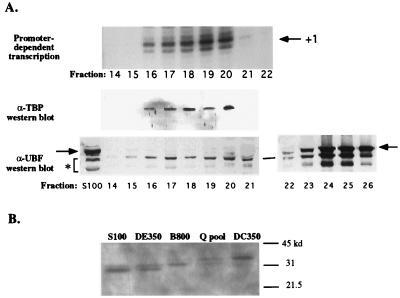
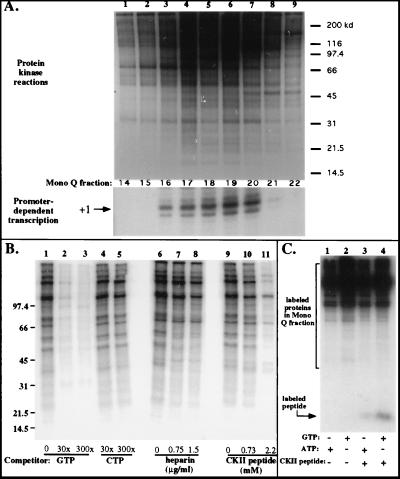
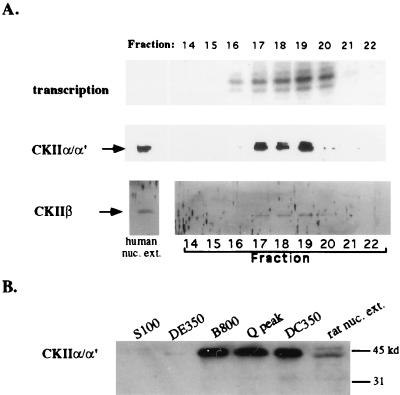
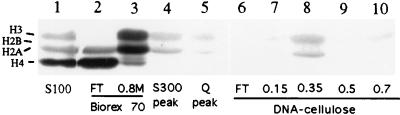
Mounting evidence suggests that eukaryotic RNA polymerases preassociate with multiple transcription factors in the absence of DNA, forming RNA polymerase holoenzyme complexes. We have purified an apparent RNA polymerase I (Pol I) holoenzyme from Xenopus laevis cells by sequential chromatography on five columns: DEAE-Sepharose, Biorex 70, Sephacryl S300, Mono Q, and DNA-cellulose. Single fractions from every column programmed accurate promoter-dependent transcription. Upon gel filtration chromatography, the Pol I holoenzyme elutes at a position overlapping the peak of Blue Dextran, suggesting a molecular mass in the range of approximately 2 MDa. Consistent with its large mass, Coomassie blue-stained sodium dodecyl sulfate-polyacrylamide gels reveal approximately 55 proteins in fractions purified to near homogeneity. Western blotting shows that TATA-binding protein precisely copurifies with holoenzyme activity, whereas the abundant Pol I transactivator upstream binding factor does not. Also copurifying with the holoenzyme are casein kinase II and a histone acetyltransferase activity with a substrate preference for histone H3. These results extend to Pol I the suggestion that signal transduction and chromatin-modifying activities are associated with eukaryotic RNA polymerases.
Figures
References
-
- Albert, A.-C. Unpublished data.
-
- Allende J E, Allende C C. Protein kinases. 4. Protein kinase CK2: an enzyme with multiple substrates and a puzzling regulation. FASEB J. 1995;9:313–323. - PubMed
-
- Bazett-Jones D, Leblanc B, Herfort M, Moss T. Short-range DNA looping by the Xenopus HMG-box transcription factor, xUBF. Science. 1994;264:1134–1137. - PubMed
-
- Bell S P, Jantzen H M, Tjian R. Assembly of alternative multiprotein complexes directs rRNA promoter selectivity. Genes Dev. 1990;4:943–954. - PubMed
-
- Bell S P, Learned R M, Jantzen H M, Tjian R. Functional cooperativity between transcription factors UBF1 and SL1 mediates human ribosomal RNA synthesis. Science. 1988;241:1192–1197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources