

Interactions between the class II transactivator and CREB binding protein increase transcription of major histocompatibility complex class II genes
- PMID: 9858618
- PMCID: PMC83952
- DOI: 10.1128/MCB.19.1.941
Interactions between the class II transactivator and CREB binding protein increase transcription of major histocompatibility complex class II genes
Abstract
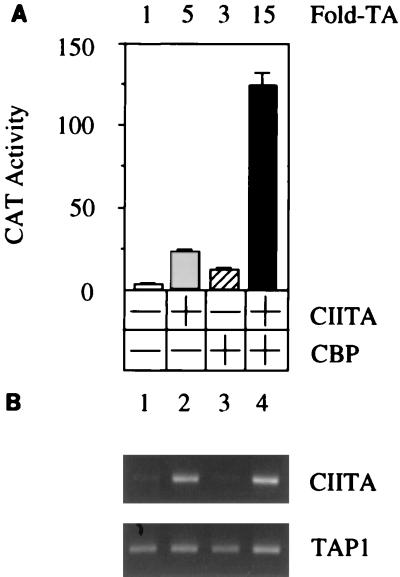
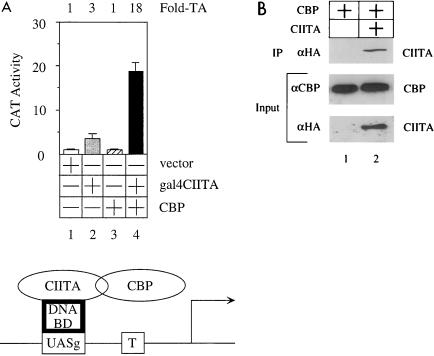
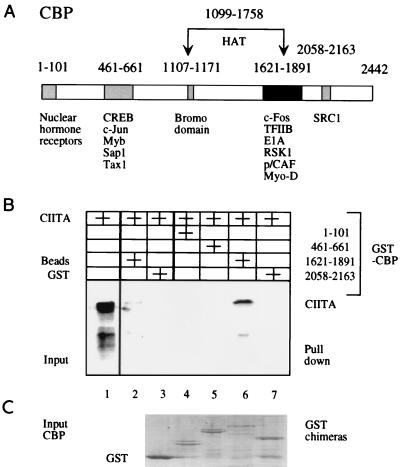
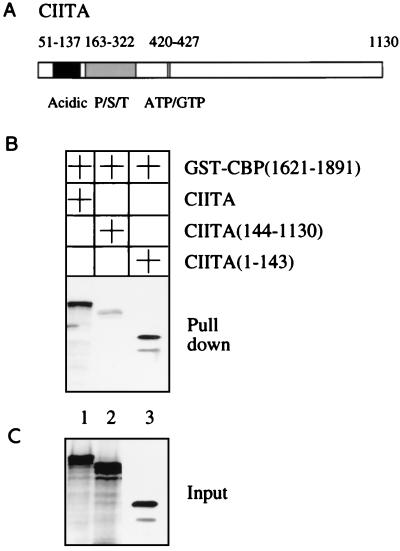
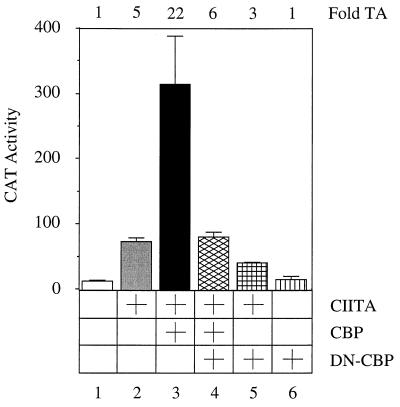
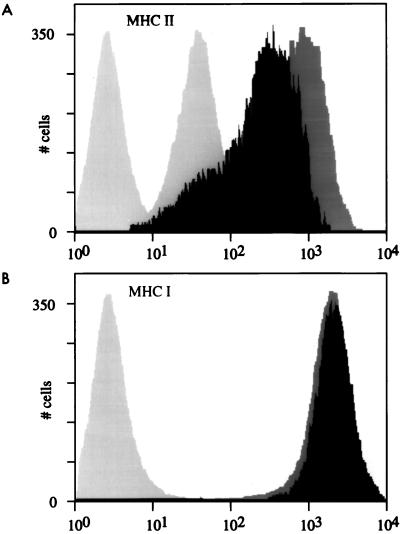
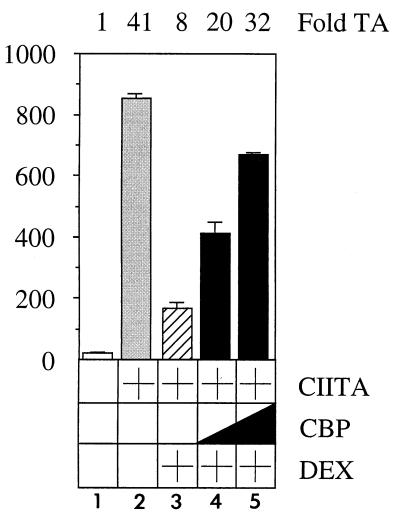
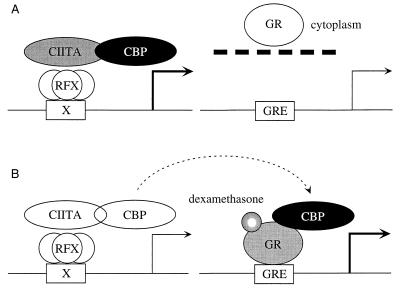
Class II major histocompatibility (class II) genes are regulated in a B-cell-specific and gamma interferon-inducible fashion. The master switch for the expression of these genes is the class II transactivator (CIITA). In this report, we demonstrate that one of the functions of CIITA is to recruit the CREB binding protein (CBP) to class II promoters. Not only functional but also specific binding interactions between CIITA and CBP were demonstrated. Moreover, a dominant negative form of CBP decreased the activity of class II promoters and levels of class II determinants on the surface of cells. Finally, the inhibition of class II gene expression by the glucocorticoid hormone could be attributed to the squelching of CBP by the glucocorticoid receptor. We conclude that CBP, a histone acetyltransferase, plays an important role in the transcription of class II genes.
Figures
References
-
- Abdulkadir S A, Ono S J. How are class II MHC genes turned on and off? FASEB J. 1995;9:1429–1435. - PubMed
-
- Abraham S E, Lobo S, Yaciuk P, Wang H G, Moran E. p300, and p300-associated proteins, are components of TATA-binding protein (TBP) complexes. Oncogene. 1993;8:1639–1647. - PubMed
-
- Arany Z, Newsome D, Oldread E, Livingston D M, Eckner R. A family of transcriptional adaptor proteins targeted by the E1A oncoprotein. Nature. 1995;374:81–84. - PubMed
-
- Arias J, Alberts A S, Brindle P, Claret F X, Smeal T, Karin M, Feramisco J, Montminy M. Activation of cAMP and mitogen responsive genes relies on a common nuclear factor. Nature. 1994;370:226–229. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources