

Gap junctional coupling in lenses lacking alpha3 connexin
- PMID: 9860964
- PMCID: PMC28038
- DOI: 10.1073/pnas.95.26.15303
Gap junctional coupling in lenses lacking alpha3 connexin
Abstract
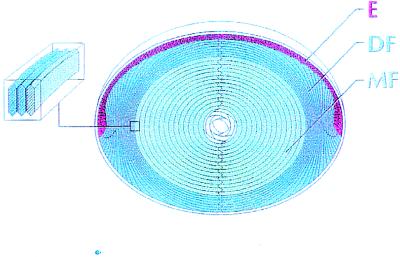
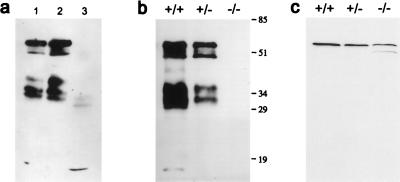
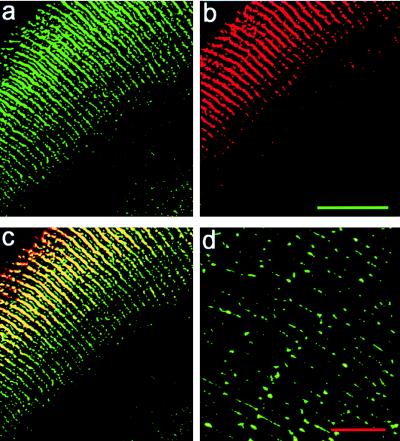
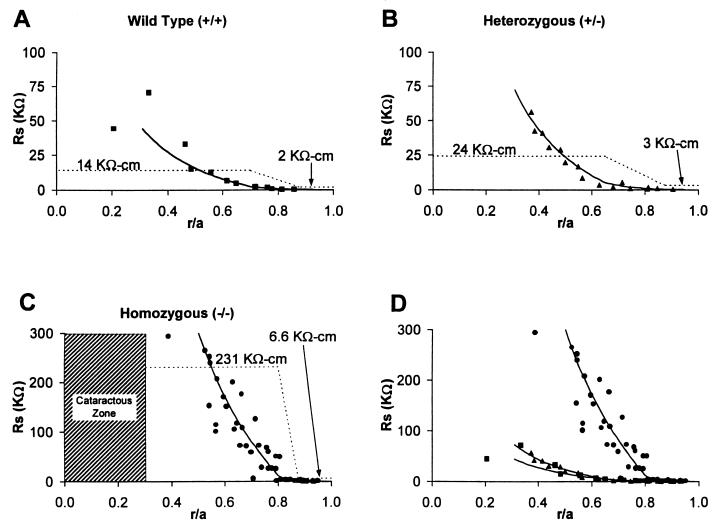
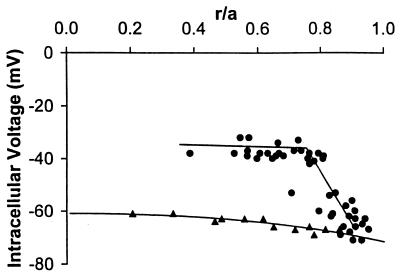
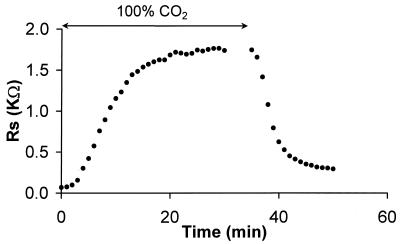
Fiber cells of the lens are interconnected by an extensive network of gap junctions containing alpha3 (Cx46) and alpha8 (Cx50) connexins. A specific role for these connexins in lens homeostasis is not known. To determine the contribution of these connexins to lens function, we used impedance techniques to study cell-to-cell coupling in lenses from homozygous alpha3 knockout (-/-), heterozygous (+/-), and wild-type (+/+) mice. Western blots and immunofluorescence data indicated that alpha8 remained at similar levels in the three classes of lenses, whereas alpha3 was approximately 50% of the normal level in the +/- lenses, and it was absent from the -/- lenses. Moreover, the data from +/+ lenses suggest that a cleavage of connexins occurs abruptly between the peripheral shell of differentiating fibers (DF) and the inner core of mature fibers (MF). The appearance of the cleaved connexins was correlated to a change in the coupling conductance. In -/- lenses the coupling conductance of MF was zero, and these fibers were depolarized by about 30 mV from normal (approximately -65 mV). The DF remained coupled, but the conductance was reduced to 30-35% of normal. However, the gap junctions in the DF of alpha3 -/- lenses remained sensitive to pH. We conclude that alpha3 connexin is necessary for the coupling of central fibers to peripheral cells, and that this coupling is essential for fiber cell homeostasis because uncoupled MF depolarize and subsequently become opaque.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
