

The cyclin-dependent kinase inhibitor p27(Kip1) induces N-terminal proteolytic cleavage of cyclin A
- PMID: 9860976
- PMCID: PMC28050
- DOI: 10.1073/pnas.95.26.15374
The cyclin-dependent kinase inhibitor p27(Kip1) induces N-terminal proteolytic cleavage of cyclin A
Abstract
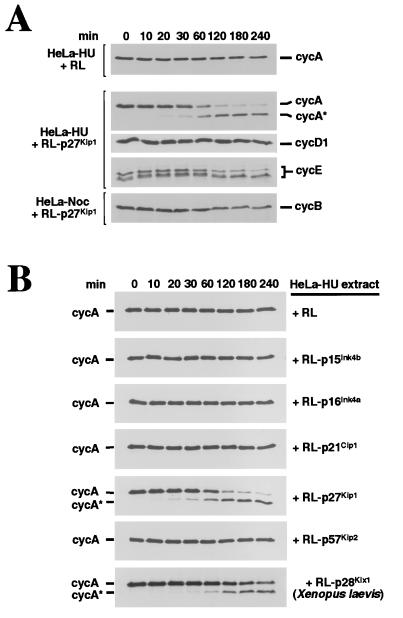
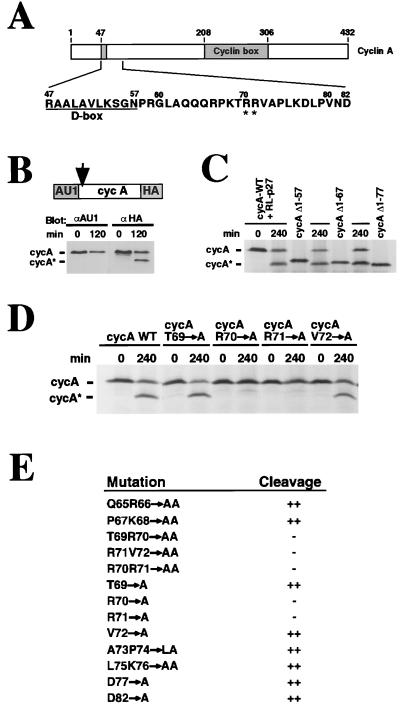
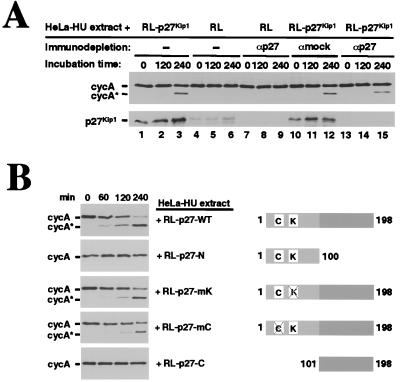
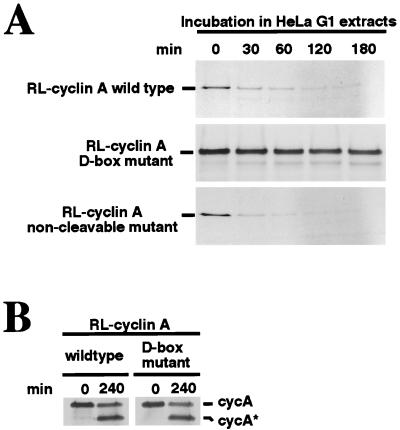
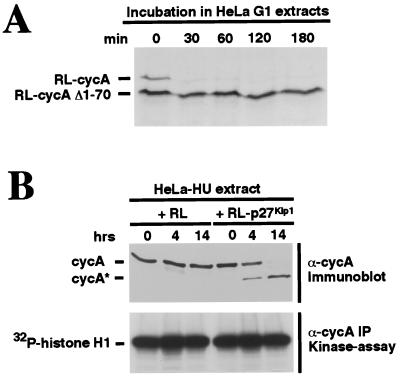
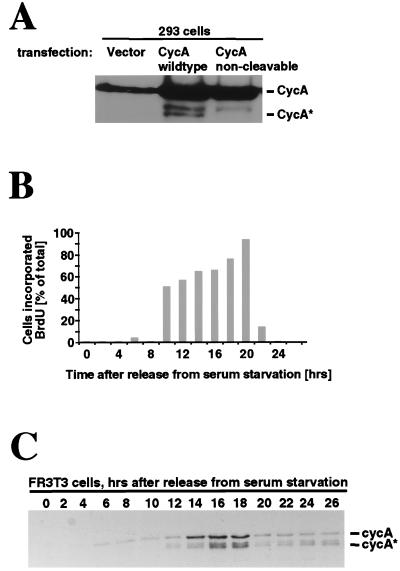
Progression through the cell cycle is regulated in part by the sequential activation and inactivation of cyclin-dependent kinases (CDKs). Many signals arrest the cell cycle through inhibition of CDKs by CDK inhibitors (CKIs). p27(Kip1) (p27) was first identified as a CKI that binds and inhibits cyclin A/CDK2 and cyclin E/CDK2 complexes in G1. Here we report that p27 has an additional property, the ability to induce a proteolytic activity that cleaves cyclin A, yielding a truncated cyclin A lacking the mitotic destruction box. Other CKIs (p15(Ink4b), p16(Ink4a), p21(Cip1), and p57(Kip2)) do not induce cleavage of cyclin A; other cyclins (cyclin B, D1, and E) are not cleaved by the p27-induced protease activity. The C-terminal half of p27, which is dispensable for its kinase inhibitory activity, is required to induce cleavage. Mechanistically, p27 does not appear to cause cleavage through direct interaction with cyclin/CDK complexes. Instead, it activates a latent protease that, once activated, does not require the continuing presence of p27. Mutation of cyclin A at R70 or R71, residues at or very close to the cleavage site, blocks cleavage. Noncleavable mutants are still recognized by the anaphase-promoting complex/cyclosome pathway responsible for ubiquitin-dependent proteolysis of mitotic cyclins, indicating that the p27-induced cleavage of cyclin A is part of a separate pathway. We refer to this protease as Tsap (pTwenty-seven- activated protease).
Figures
References
-
- Sherr C J. Cell. 1993;73:1059–1065. - PubMed
-
- Pines J. Trends Biochem Sci. 1993;18:195–197. - PubMed
-
- Hengst L, Reed S I. Curr Top Microbiol Immunol. 1998;227:25–41. - PubMed
-
- Sherr C, Roberts J. Genes Dev. 1995;9:1149–1163. - PubMed
-
- King R W, Deshaies R J, Peters J, Kirschner M. Science. 1996;274:1652–1659. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
