

RNA as a target of double-stranded RNA-mediated genetic interference in Caenorhabditis elegans
- PMID: 9860998
- PMCID: PMC28072
- DOI: 10.1073/pnas.95.26.15502
RNA as a target of double-stranded RNA-mediated genetic interference in Caenorhabditis elegans
Abstract
Introduction of exogenous double-stranded RNA (dsRNA) into Caenorhabditis elegans has been shown to specifically and potently disrupt the activity of genes containing homologous sequences. In this study we present evidence that the primary interference effects of dsRNA are post-transcriptional. First, we examined the primary DNA sequence after dsRNA-mediated interference and found no evidence for alterations. Second, we found that dsRNA-mediated interference with the upstream gene in a polar operon had no effect on the activity of the downstream gene; this finding argues against an effect on initiation or elongation of transcription. Third, we observed by in situ hybridization that dsRNA-mediated interference produced a substantial, although not complete, reduction in accumulation of nascent transcripts in the nucleus, while cytoplasmic accumulation of transcripts was virtually eliminated. These results indicate that the endogenous mRNA is the target for interference and suggest a mechanism that degrades the targeted RNA before translation can occur. This mechanism is not dependent on the SMG system, an mRNA surveillance system in C. elegans responsible for targeting and destroying aberrant messages. We suggest a model of how dsRNA might function in a catalytic mechanism to target homologous mRNAs for degradation.
Figures
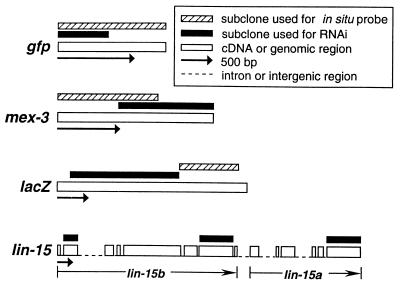
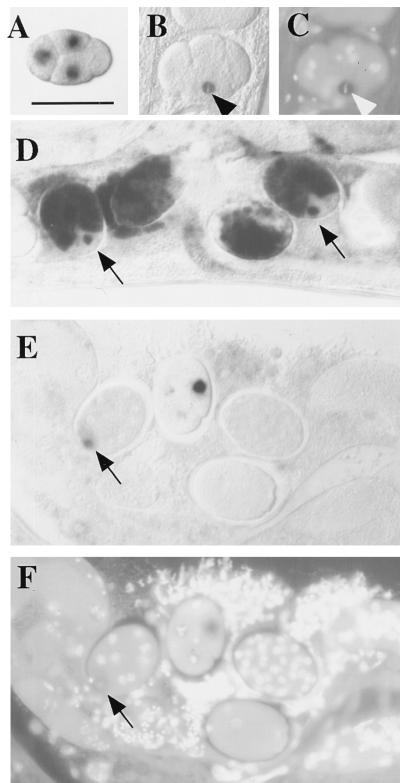
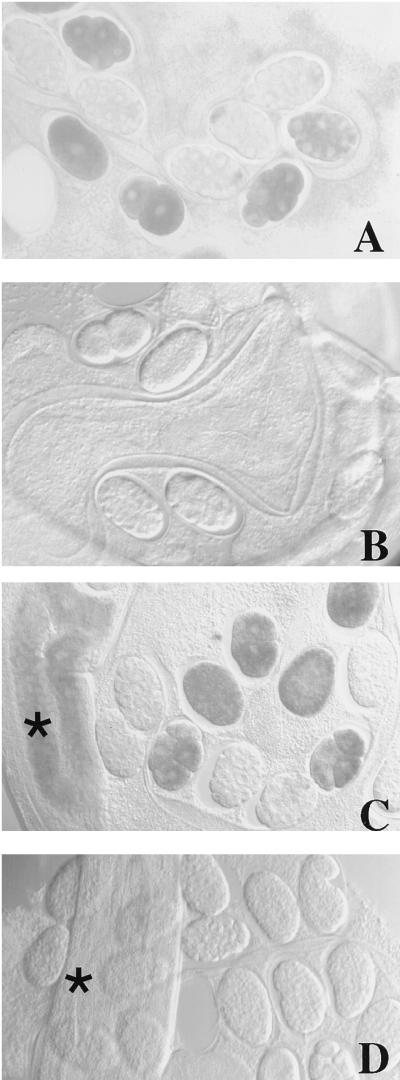
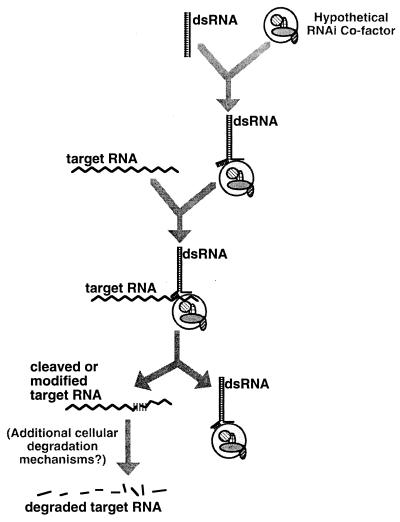
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
