

The Arabidopsis thaliana RPM1 disease resistance gene product is a peripheral plasma membrane protein that is degraded coincident with the hypersensitive response
- PMID: 9861059
- PMCID: PMC28133
- DOI: 10.1073/pnas.95.26.15849
The Arabidopsis thaliana RPM1 disease resistance gene product is a peripheral plasma membrane protein that is degraded coincident with the hypersensitive response
Abstract
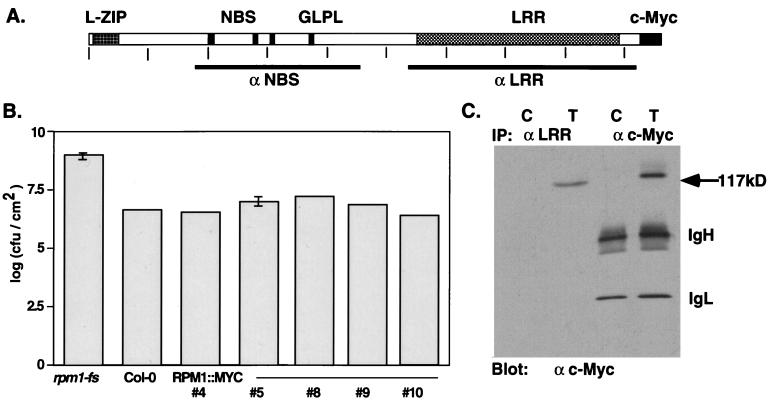
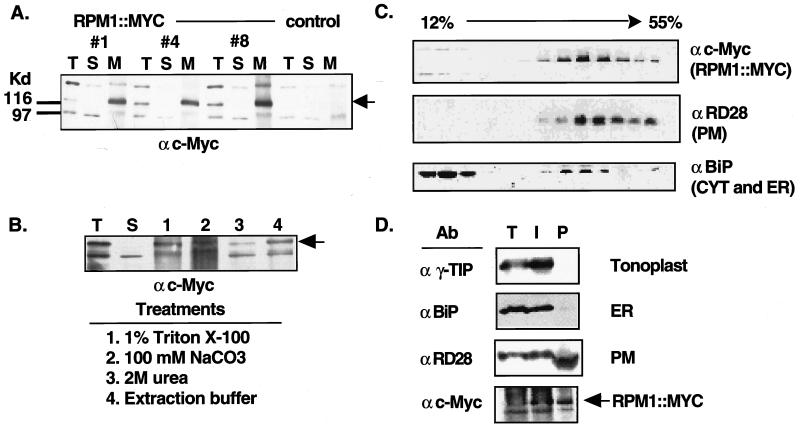
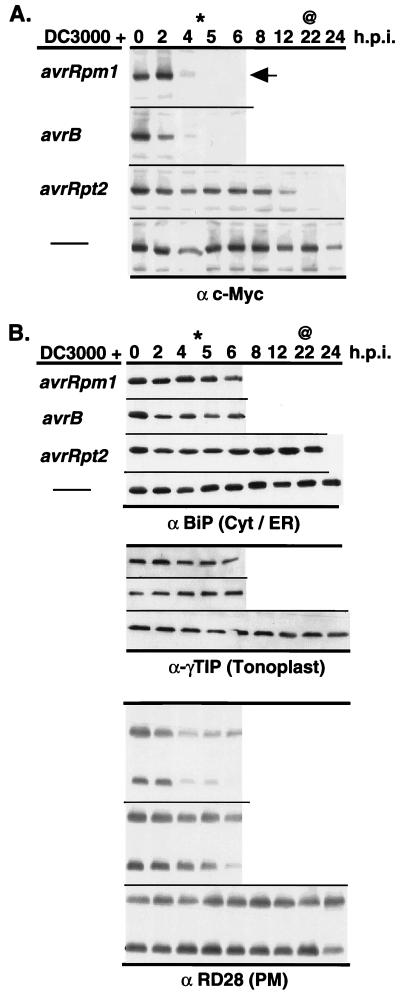
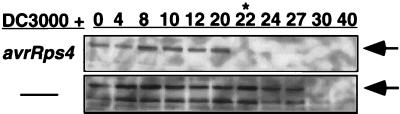
Disease resistance in plants is often controlled by a gene-for-gene mechanism in which avirulence (avr) gene products encoded by pathogens are specifically recognized, either directly or indirectly, by plant disease resistance (R) gene products. Members of the NBS-LRR class of R genes encode proteins containing a putative nucleotide binding site (NBS) and carboxyl-terminal leucine-rich repeats (LRRs). Generally, NBS-LRR proteins do not contain predicted transmembrane segments or signal peptides, suggesting they are soluble cytoplasmic proteins. RPM1 is an NBS-LRR protein from Arabidopsis thaliana that confers resistance to Pseudomonas syringae expressing either avrRpm1 or avrB. RPM1 protein was localized by using an epitope tag. In contrast to previous suggestions, RPM1 is a peripheral membrane protein that likely resides on the cytoplasmic face of the plasma membrane. Furthermore, RPM1 is degraded coincident with the onset of the hypersensitive response, suggesting a negative feedback loop controlling the extent of cell death and overall resistance response at the site of infection.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
