

Interaction of NK lysin, a peptide produced by cytolytic lymphocytes, with endotoxin
- PMID: 9864216
- PMCID: PMC96297
- DOI: 10.1128/IAI.67.1.201-205.1999
Interaction of NK lysin, a peptide produced by cytolytic lymphocytes, with endotoxin
Abstract
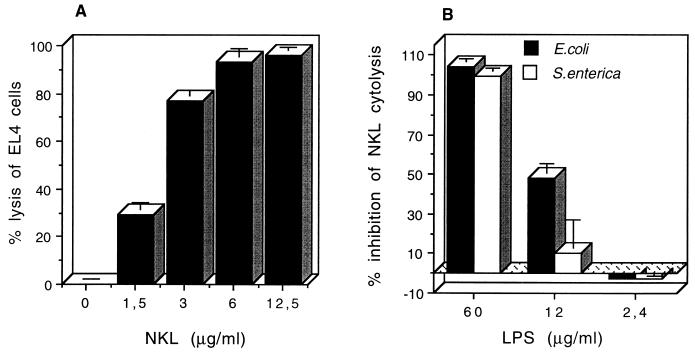
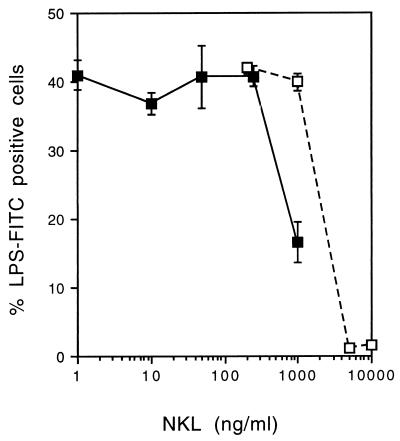
NK lysin is a 9-kDa polypeptide that was originally isolated from porcine intestinal tissue based on its antibacterial activity. It is produced by cytolytic lymphocytes and is cytolytic against a number of different types of tumor cells. Here we report the binding of NK lysin to lipopolysaccharide (LPS) and its anti-LPS activity. NK lysin binds to matrix-coated LPS from Escherichia coli, Pseudomonas aeruginosa, and different strains of Salmonella enterica. Lipid A and polymyxin B inhibited the binding, demonstrating a preferential interaction of NK lysin with the lipid part of LPS. Chromium-labeled lymphoma cells were lysed by NK lysin, and LPS dose-dependently inhibited the cytolysis at equimolar amounts. In the same manner, NK lysin inhibited certain LPS-stimulated effects on mouse bone marrow cells as well as LPS binding to mouse granulocytes. These results suggest that NK lysin may be a another natural LPS-binding protein from lymphocytes that may participate in the endogenous defense response associated with elevated concentrations of LPS.
Figures

References
-
- Andersson M, Curstedt T, Jörnvall H, Johansson J. An amphipathic helical motif common to tumourolytic polypeptide NK-lysin and pulmonary surfactant polypeptide SP-B. FEBS Lett. 1995;362:328–332. - PubMed
-
- Andersson M, Gunne H, Agerberth B, Boman A, Bergman T, Sillard R, Jörnvall H, Mutt V, Olsson B, Wigzell H, Dagerlind Å, Boman G H, Gudmundsson H G. NK-lysin, a novel effector peptide of cytotoxic T and NK cells. Structure and cDNA cloning of the porcine form, induction by interleukin 2, antibacterial and antitumour activity. EMBO J. 1995;14:1615–1625. - PMC - PubMed
-
- Andersson, M., and B. Olsson. Unpublished data.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
