

Gamma interferon augments macrophage activation by lipopolysaccharide by two distinct mechanisms, at the signal transduction level and via an autocrine mechanism involving tumor necrosis factor alpha and interleukin-1
- PMID: 9864217
- PMCID: PMC96298
- DOI: 10.1128/IAI.67.1.206-212.1999
Gamma interferon augments macrophage activation by lipopolysaccharide by two distinct mechanisms, at the signal transduction level and via an autocrine mechanism involving tumor necrosis factor alpha and interleukin-1
Abstract
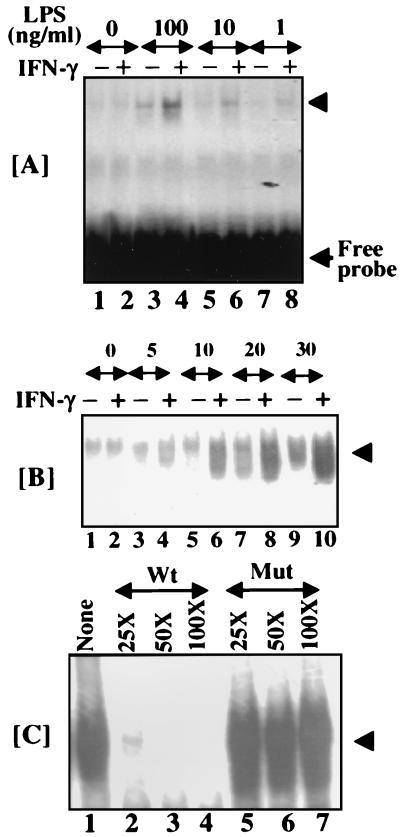
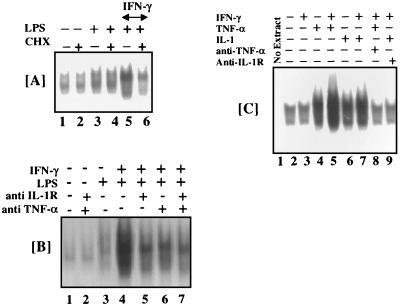
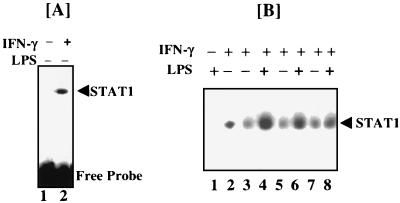
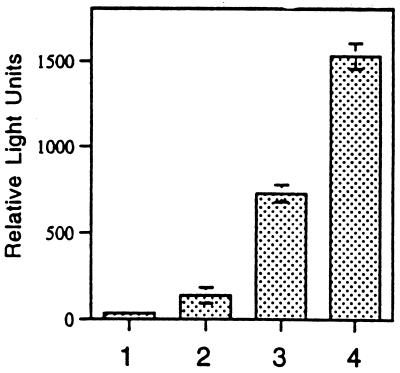
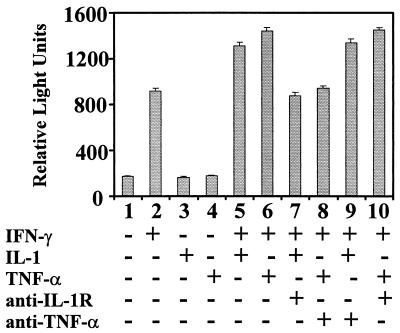
When given in the presence of gamma interferon (IFN-gamma), otherwise nontoxic doses of lipopolysaccharide (LPS or endotoxin) become highly lethal for mice. The mechanisms of this synergistic toxicity are not known. We considered the possibility that an interaction between the LPS-induced NF-kappaB and IFN-gamma-induced JAK-STAT pathways at the pretranscriptional level may enhance the LPS-induced signals. To test this hypothesis, we incubated murine macrophage RAW 264.7 cells with IFN-gamma for 2 h before addition of different doses of LPS. Consistent with the synergistic induction of inducible nitric oxide synthase mRNA and nitric oxide production by a combination of LPS and IFN-gamma, IFN-gamma strongly augmented LPS-induced NF-kappaB activation and accelerated the binding of NF-kappaB to DNA to as early as 5 min. In agreement with this, IFN-gamma pretreatment promoted rapid degradation of IkappaB-alpha but not that of IkappaB-beta. Inhibition of protein synthesis during IFN-gamma treatment suppressed LPS-initiated NF-kappaB binding. A rapidly induced protein appeared to be involved in IFN-gamma priming. Preincubation of cells with antibodies to tumor necrosis factor alpha or the interleukin-1 receptor partially reduced the priming effect of IFN-gamma. In a complementary manner, LPS enhanced the activation of signal-transducing activator of transcription 1 by IFN-gamma. These data suggest novel mechanisms for the synergy between IFN-gamma and LPS by which they cross-regulate the signal-transducing molecules. Through this mechanism, IFN-gamma may transform a given dose of LPS into a lethal stimulus capable of causing sepsis. It may also serve a beneficial purpose by enabling the host to respond quickly to relatively low doses of LPS and thereby activating antibacterial defenses.
Figures

References
-
- Ausubel F M, Brent R, Kingston D D, Moore J G, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. New York, N.Y: Wiley-Interscience; 1994.
-
- Baeuerle P A, Henkel T. Function and activation of NF-kappa B in the immune system. Annu Rev Immunol. 1994;12:141–179. - PubMed
-
- Bonizzi G, Dejardin E, Piret B, Piette J, Merville M P, Bours V. Interleukin-1 beta induces nuclear factor kappa B in epithelial cells independently of the production of reactive oxygen intermediates. Eur J Biochem. 1996;242:544–549. - PubMed
-
- Cannon J G, Tompkins R G, Gelfand J A, Michie H R, Stanford G G, van der Meer J W, Endres S, Lonnemann G, Corsetti J, Chernow B, et al. Circulating interleukin-1 and tumor necrosis factor in septic shock and experimental endotoxin fever. J Infect Dis. 1990;161:79–84. - PubMed
-
- Crofford L J, Tan B, McCarthy C J, Hla T. Involvement of nuclear factor kappa B in the regulation of cyclooxygenase-2 expression by interleukin-1 in rheumatoid synoviocytes. Arthritis Rheum. 1997;40:226–236. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
