

doi: 10.1101/gad.12.24.3815.
Molecular interactions between Vestigial and Scalloped promote wing formation in Drosophila
Affiliations
- PMID: 9869635
- PMCID: PMC317270
- DOI: 10.1101/gad.12.24.3815
Item in Clipboard
Molecular interactions between Vestigial and Scalloped promote wing formation in Drosophila
Genes Dev.
.
Abstract
Scalloped (Sd) and Vestigial (Vg) are each needed for Drosophila wing development. We show that Sd is required for Vg function and that altering their relative cellular levels inhibits wing formation. In vitro, Vg binds directly to both Sd and its human homolog, Transcription Enhancer Factor-1. The interaction domains map to a small region of Vg that is essential for Vg-mediated gene activation and to the carboxy-terminal half of Sd. Our observations indicate that Vg and Sd function coordinately to control the expression of genes required for wing development, which implies that Vg is a tissue-specific transcriptional intermediary factor of Sd.
Figures
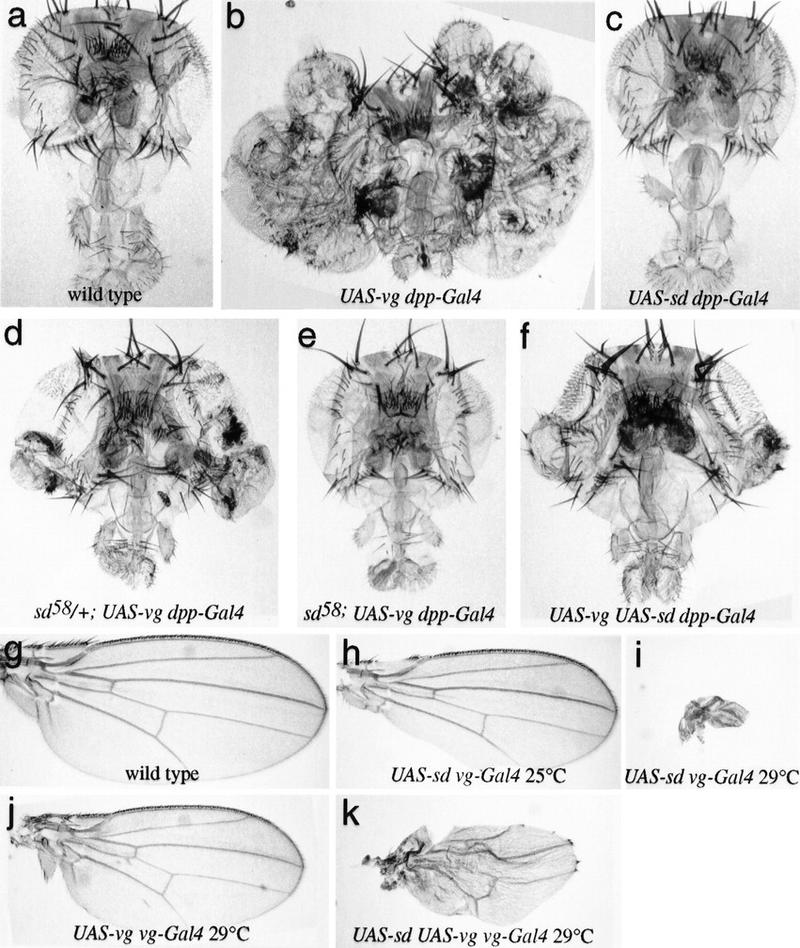
Vg requires Sd and is sensitive to Sd levels in vivo. (a–f) Drosphila heads; (g–k) Drosphila wings, in which the Vg and/or Sd proteins have been ectopically expressed using the UAS–Gal4 system (Brand and Perrimon 1993). (a) Wild type. (b) UAS–vg dpp–GAL4. A massive outgrowth of wing tissue from the eye occurs. (c) UAS–sd dpp–GAL4. No wing tissue is induced, and higher levels of expression (e.g., using ptc–GAL4) result in loss of head tissue. (d) sd58/+; UAS–vg dpp–GAL4. The sd58 mutation reduces the amount of Sd produced, and formation of ectopic wing tissue is partially suppressed. (e) sd58; UAS–vg dpp–GAL4. The formation of wing tissue is completely suppressed when no wild-type copy of sd is present. (f) UAS–vg UAS–sd dpp–GAL4. The formation of wing tissue is also partially suppressed when levels of Sd are increased. Raising or lowering the levels of Sd relative to that of Vg within the developing wing disc produces corresponding phenotypes. (g) Wild type. (h) UAS–sd vg–GAL4 at 25°C. Elevating Sd levels in wing progenitor cells leads to incomplete formation of the wing margin. (i) UAS–sd vg–GAL4 at 29°C. This effect becomes more severe in flies raised at 29°C, where almost no wing tissue is formed. (j) UAS–vg vg–GAL4 at 29°C. The elevated Vg levels in the wings of these animals leads to a slight reduction in wing size. (k) UAS–sd UAS–vg vg–GAL4 at 29°C. The loss of wing tissue induced by excess Sd (i) is partially suppressed by the simultaneous increase in Vg expression.
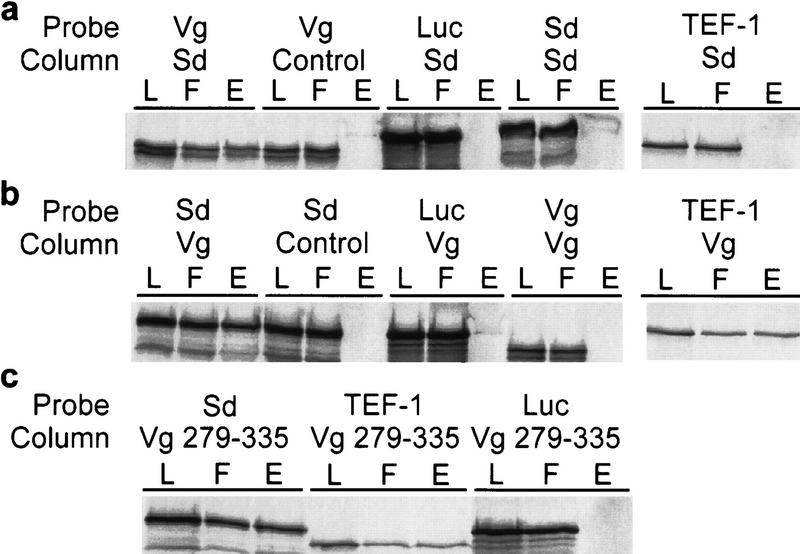
Vg and Sd exhibit a strong protein–protein interaction in vitro. (a) Affinity chromatography was performed using bound bacterially expressed Sd protein [(L) column load; (F) column flowthrough; (E) column eluate]. Sd columns selectively retain Vg and not Sd or TEF-1. As a control for nonspecific binding, Luc was expressed in the same in vitro system and shown to not bind to the Sd column. Similarly, the column matrix alone does not retain significant amounts of labeled Vg. (b) Vg columns specifically bind Sd protein but not Vg or Luc. The human TEF-1 protein, a homolog of Drosphila Sd, also shows a similar specific affinity for Vg. (c) Affinity columns bound with a protein consisting of Vg amino acids 279–335 (see Fig. 3) can selectively retain Sd or TEF-1 but not Luc.
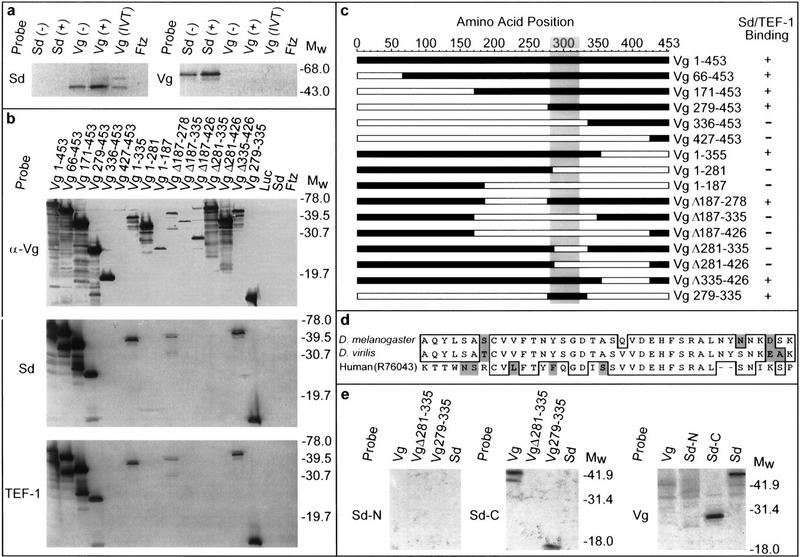
Mapping the Vg–Sd interaction domain. (a) Far Western blots of bacterial or in vitro-translated (IVT) Vg or Sd were probed with labeled Vg or Sd probes. Ftz protein expressed under similar conditions was also included on each blot to detect nonspecific probe binding. Sd selectively binds bacterially produced and IVT Vg, whereas a corresponding blot probed with Vg shows binding only to Sd. As the detected band is more intense in cell extracts after induction (+) of the expression plasmid than before (−), this confirms that each is plasmid and not endogenously encoded. (b) All but one of the specific deletions of Vg (c) were recognized by anti-Vg antibody (α-Vg), confirming that Vg is present in each lane. Expression of the Vg 427–453 protein was verified by Coomassie staining. When these deleted proteins were probed with Sd or TEF-1 under conditions similar to those above, only proteins retaining amino acids 279–335 of Vg were detected. Luc, Sd, or Ftz control lanes did not show any signals. (c) A map summarizing the position and relative size of the deletions (open bars) within Vg assayed for binding (+ or −) to Sd or TEF-1. The shaded area denotes a region that is highly similar to Vg from D. virilis and A. aegypti (S. Carroll, pers. comm.). (d) This region is also highly similar to sequences identified in mammalian genomic and expressed sequence tag databases (GenBank/EST accession nos. Z798880, Z97632, AA474871, AA571483, W81241, R65857, R73306, R76043). In addition to the 38 amino acids indicated, the smallest tested portion of Vg that has Sd-binding ability includes an amino-terminal Q and amino acids (ESSSPMSSRNFPPSFWN) carboxy-terminal to this homologous region. (e) Probes corresponding to the amino-terminal half of Sd (amino acids 1–242, Sd-N) show no interaction with Vg. The carboxy-terminal half of Sd (amino acids 240–440, Sd-C) binds to Vg protein containing only amino acids 279–335 (Sd-binding domain). A reciprocal blot using full-length Vg (Vg) as a probe shows interaction only with the carboxy-terminal half of Sd (Sd-C).
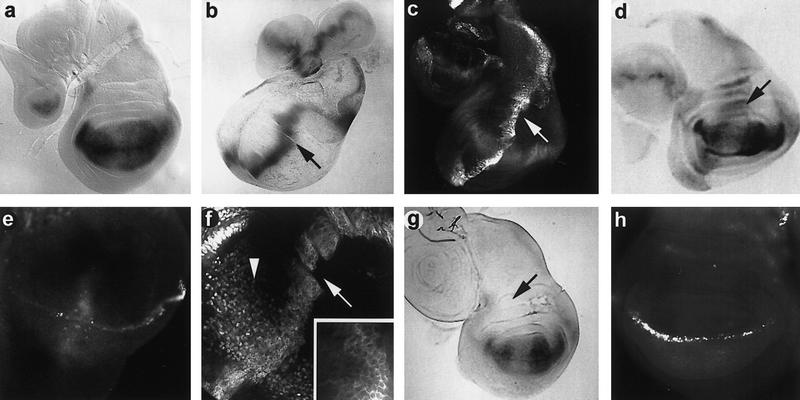
Influence of ectopic Vg on gene expression. (a) Third instar larval wing discs of genotype sd–lacZ; ptc–GAL4 showing a normal pattern of high levels of sd–lacZ expression in all cells of the presumptive wing pouch (Campbell et al. 1992). Using in situ hybridization to sd mRNA, low levels of sd expression are also detected outside of the wing pouch (not shown). A localized depression of sd activity is usually seen where the D/V intersects the A/P boundary. (b) ptc–GAL4 activates a UAS–lacZ reporter along the A/P axis of the wing disc. Note that the highest levels of activation occur in cells immediately adjacent to the border (arrow); lower levels of lacZ expression are seen in more anterior cells. (c) A ptc–GAL4; UAS–vg wing imaginal disc shows a similar graded pattern of ectopic vg expression along the A/P margin (arrow). (d) Third instar wing discs of genotype sd–lacZ; ptc–GAL4; UAS–vg. The ectopic vg expression activates sd along the A/P margin, causing a nongraded level of activation of the sd–lacZ reporter gene in vg-expressing cells (arrow). (e) A wing imaginal disc of the same genotype as c stained with anti-ct antibody. Ectopic activation of ct occurs in some cells that express ectopic Vg and Sd along the A/P margin. (f) VgΔ281–335 protein (driven by ptc–GAL4) is found primarily in the cytoplasm [arrow and inset (high magnification)] compared to the nuclear localization of endogenous wild-type Vg (arrowhead). (g) In discs containing sd–lacZ, ptc–GAL4 mediated expression of a UAS–vgΔ281–335 construct (missing the Sd-binding domain) does not induce sd expression along the A/P margin (arrow). (h) Where high levels of ectopic VgΔ281–335 and Sd overlap, ct expression remains in the wild-type pattern along the D/V border.
References
-
- Blair SS. A role for the segment polarity gene shaggy-zeste white 3 in the specification of regional identity in the developing wing of Drosphila. Dev Biol. 1994;162:229–244. - PubMed
-
- Blochlinger KR, Bodmer R, Jan LY, Jan YN. Patterns of expression of cut, a protein required for external sensory organ development in wild type and mutant Drosphila embryos. Genes & Dev. 1990;4:1322–1331. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Campbell SD, Inamdar M, Rodrigues V, Raghavan V, Palazzolo M, Chovnick A. The scalloped gene encodes a novel, evolutionarily conserved transcription factor required for sensory organ differentiation in Drosphila. Genes & Dev. 1992;6:367–379. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases