

The Vestigial and Scalloped proteins act together to directly regulate wing-specific gene expression in Drosophila
- PMID: 9869643
- PMCID: PMC317267
- DOI: 10.1101/gad.12.24.3900
The Vestigial and Scalloped proteins act together to directly regulate wing-specific gene expression in Drosophila
Abstract
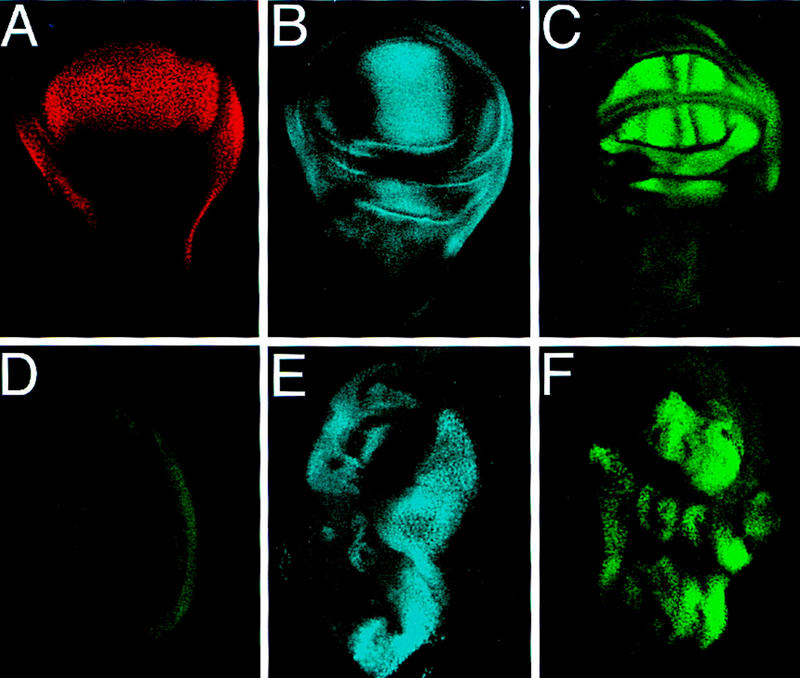
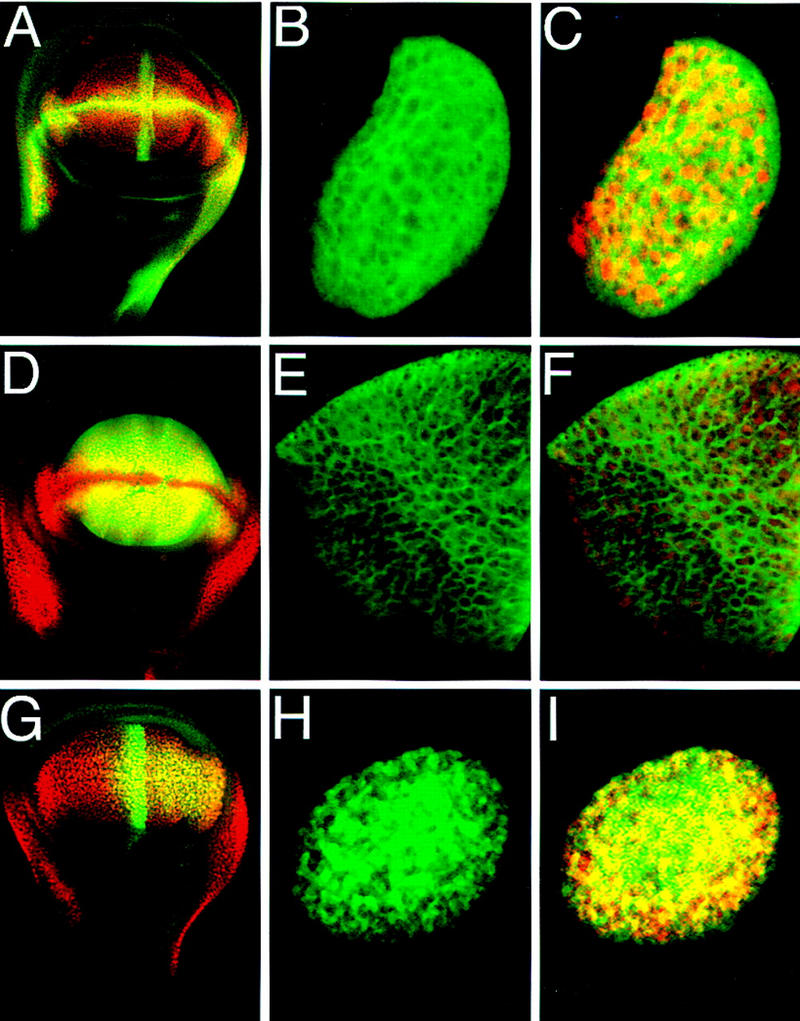
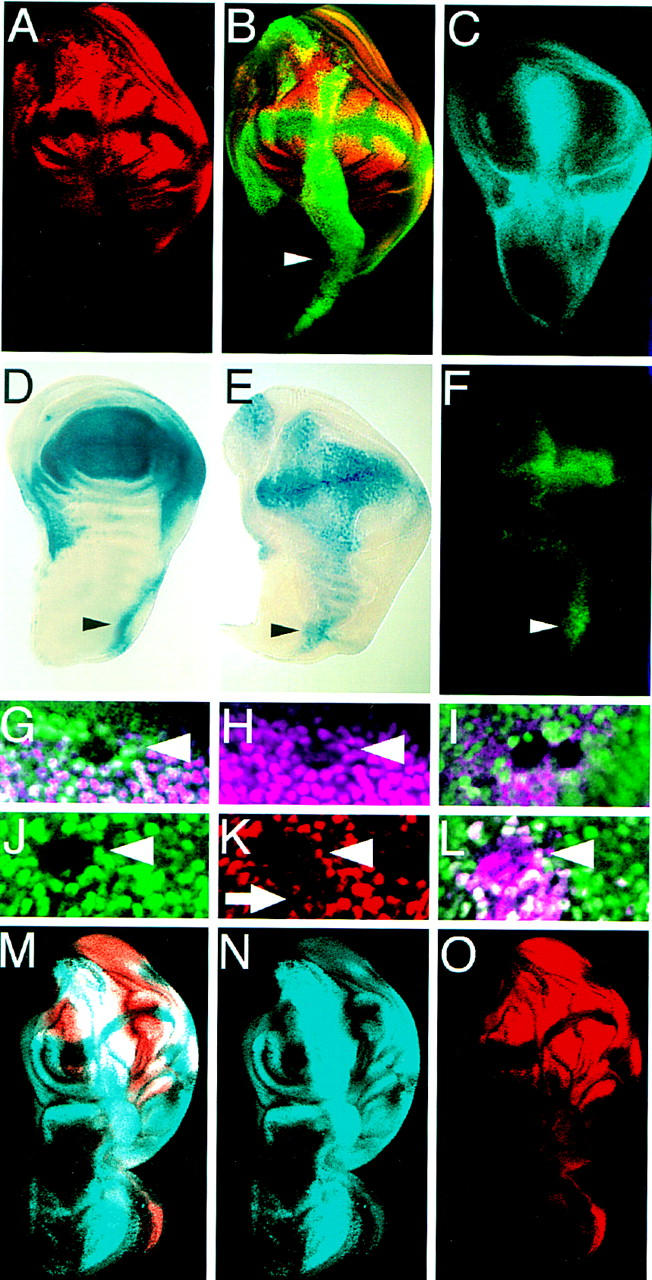
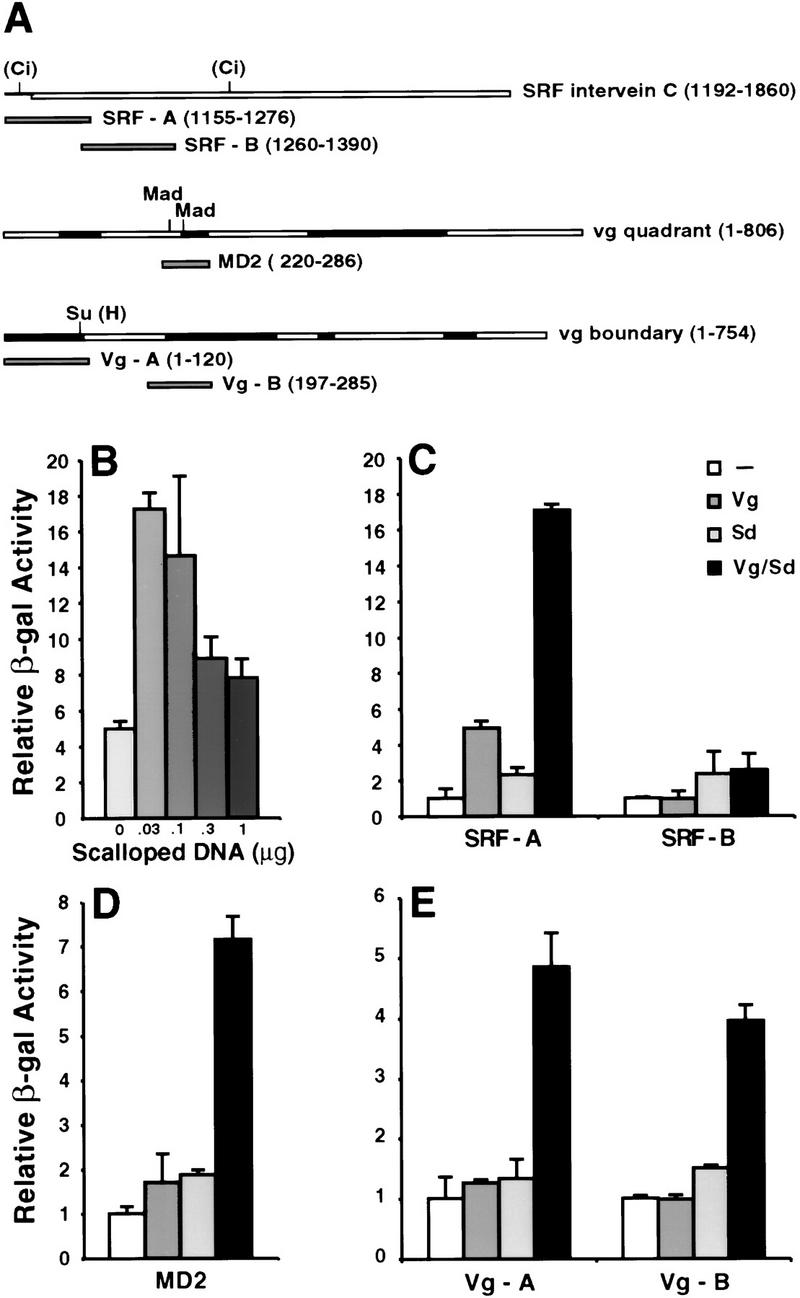
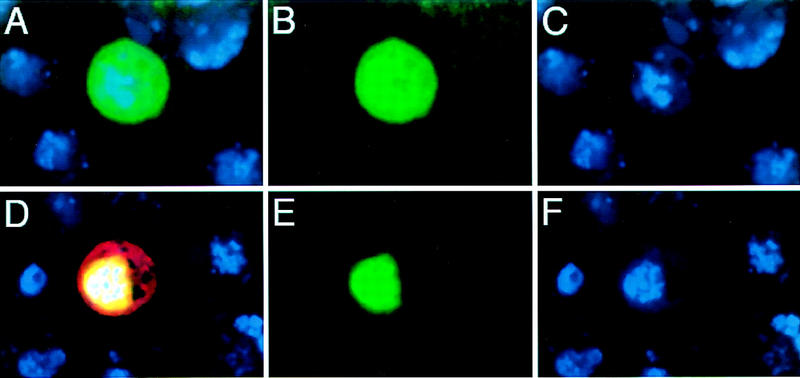
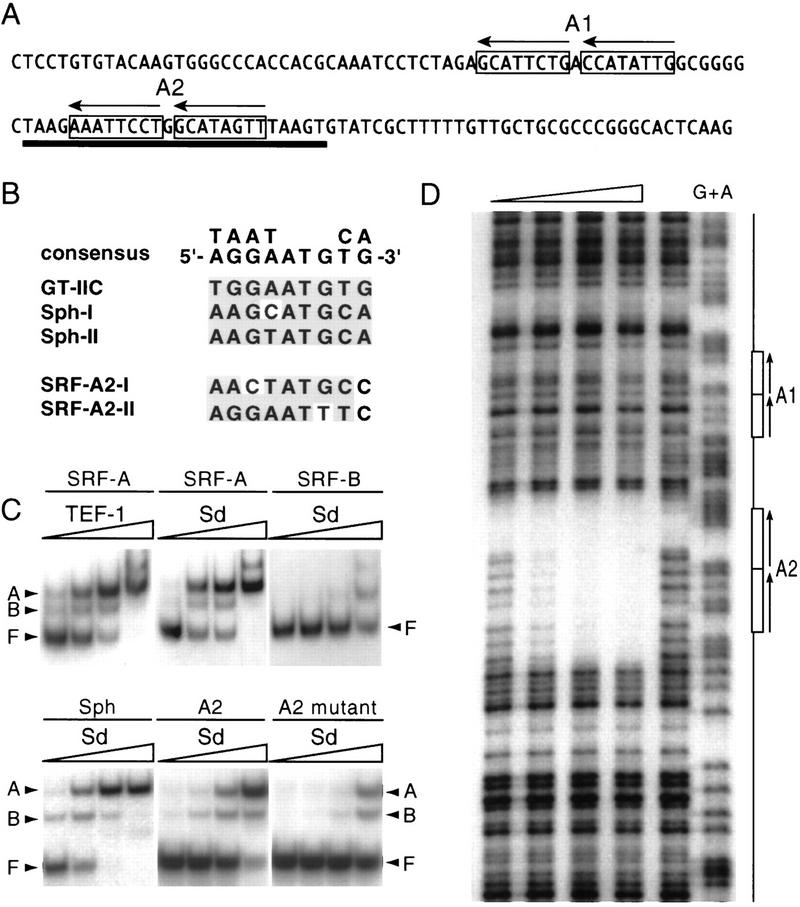
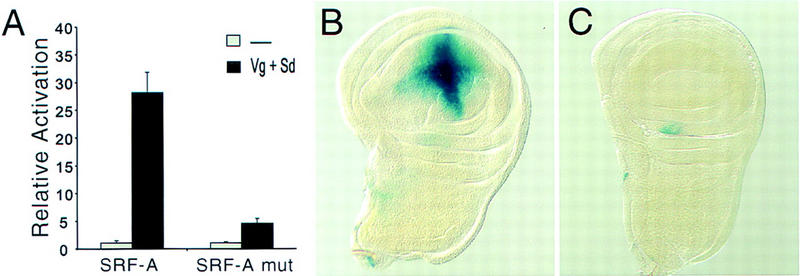
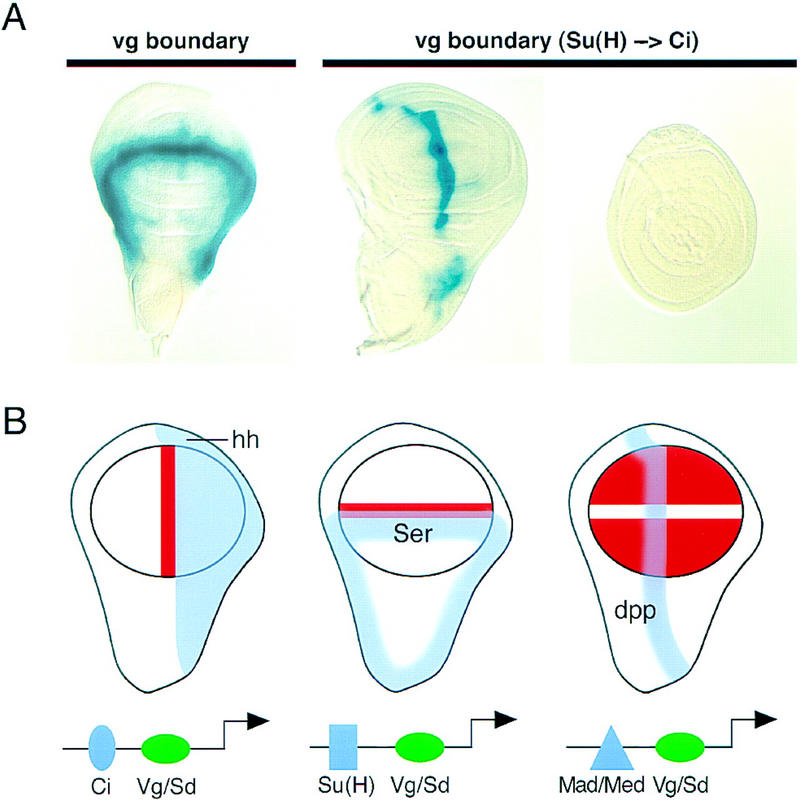
A small number of major regulatory (selector) genes have been identified in animals that control the development of particular organs or complex structures. In Drosophila, the vestigial gene is required for wing formation and is able to induce wing-like outgrowths on other structures. However, the molecular function of the nuclear Vestigial protein, which bears no informative similarities to other proteins, was unknown. Here, we show that Vestigial requires the function of the Scalloped protein, a member of the TEA family of transcriptional regulators, to directly activate the expression of genes involved in wing morphogenesis. Genetic and molecular analyses reveal that Vestigial regulates wing identity by forming a complex with the Scalloped protein that binds sequence specifically to essential sites in wing-specific enhancers. These enhancers also require the direct inputs of signaling pathways, and the response of an enhancer can be switched to another pathway through changes in signal-transducer binding sites. Combinatorial regulation by selector proteins and signal transducers is likely to be a general feature of the tissue-specific control of gene expression during organogenesis.
Figures
References
-
- Bailey A, Posakony JW. Suppressor of hairless directly activates transcription of enhancer of split complex genes in response to Notch receptor activity. Genes & Dev. 1995;9:2609–2622. - PubMed
-
- Brand A, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Brook W, Diaz-Benjumea F, Cohen S. Organizing spatial pattern in limb development. Annu Rev Cell Dev Biol. 1996;12:161–180. - PubMed
-
- Calleja M, Moreno E, Pelaz S, Morata G. Visualization of gene expression in living adult Drosophila. Science. 1996;274:252–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases