

Mechanisms and structural determinants of HIV-1 coat protein, gp41-induced neurotoxicity
- PMID: 9870939
- PMCID: PMC6782354
- DOI: 10.1523/JNEUROSCI.19-01-00064.1999
Mechanisms and structural determinants of HIV-1 coat protein, gp41-induced neurotoxicity
Abstract
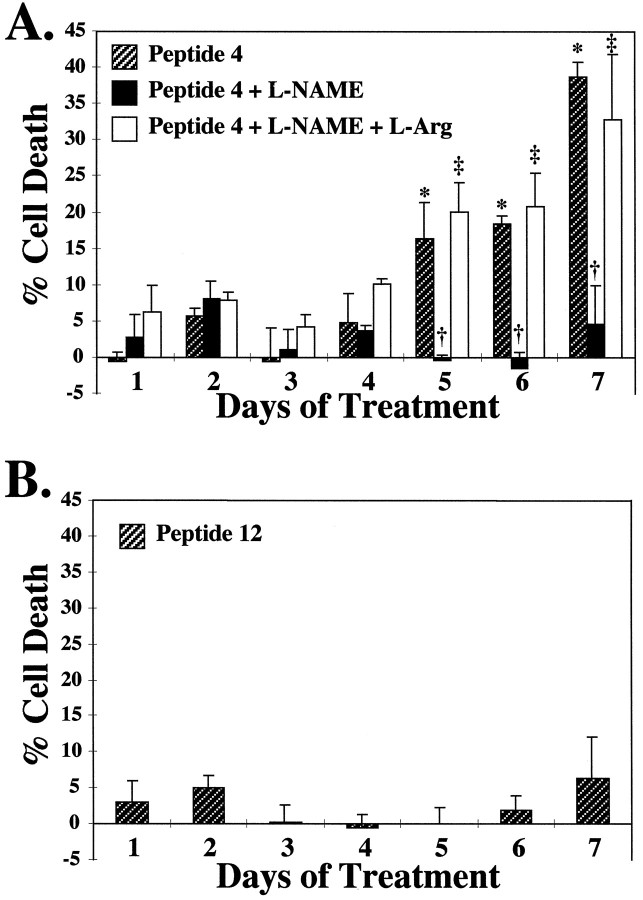
Of the individuals with human immunodeficiency virus type 1 (HIV-1) infection, 20-30% will develop the neurological complication of HIV-associated dementia (HAD). The mechanisms underlying HAD are unknown; however, indirect immunologically mediated mechanisms are theorized to play a role. Recently, the HIV-1 coat protein gp41 has been implicated as a major mediator of HAD through induction of neurocytokines and subsequent neuronal cell death. Using primary mixed cortical cultures from neuronal nitric oxide synthase (NOS) null (nNOS-/-) mice and immunological NOS null (iNOS-/-) mice, we establish iNOS-derived NO as a major mediator of gp41 neurotoxicity. Neurotoxicity elicited by gp41 is markedly attenuated in iNOS-/- cultures compared with wild-type and nNOS-/- cultures. The NOS inhibitor L-nitroarginine methyl ester is neuroprotective in wild-type and nNOS-/- cultures, confirming the role of iNOS-derived NO in gp41 neurotoxicity. Confirming that iNOS-/- cultures lack iNOS, gp41 did not induce iNOS in iNOS-/- cultures, but it markedly induced iNOS in wild-type and nNOS-/- cultures. We elucidate the region of gp41 that is critical for iNOS induction and neuronal cell death by monitoring iNOS induction with overlapping peptides spanning gp41. We show that the N-terminal region of gp41, which we designate as the neurotoxic domain, induces iNOS protein activity and iNOS-dependent neurotoxicity at picomolar concentrations in a manner similar to recombinant gp41 protein. Our experiments suggest that gp41 is eliciting the induction of iNOS through potential cell surface receptors or binding sites because the induction of iNOS is dose dependent and saturable and occurs at physiologically relevant concentrations. These data confirm that the induction of iNOS by gp41 and the production of NO are primary mediators of neuronal damage and identify a neurotoxic domain of gp41 that may play an important role in HAD.
Figures






References
-
- Adamson DC, Wildemann B, Sasaki M, Glass JD, McArthur JC, Christov VI, Dawson TM, Dawson VL. Immunologic NO synthase: elevation in severe AIDS dementia and induction by HIV-1 coat protein, gp41. Science. 1996b;274:1917–1921. - PubMed
-
- Alkhatib G, Combadiere C, Broder CC, Feng Y, Kennedy PE, Murphy PM, Berger EA. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272:1955–1958. - PubMed
-
- Barcova M, Kacani L, Speth C, Dierich MP. gp41 envelope protein of human immunodeficiency virus induces interleukin (IL)-10 in monocytes, but not in B, T, or NK cells, leading to reduced IL-2 and interferon-gamma production. J Infect Dis. 1998;177:905–913. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources