

Effects of attention on orientation-tuning functions of single neurons in macaque cortical area V4
- PMID: 9870971
- PMCID: PMC6782389
- DOI: 10.1523/JNEUROSCI.19-01-00431.1999
Effects of attention on orientation-tuning functions of single neurons in macaque cortical area V4
Abstract
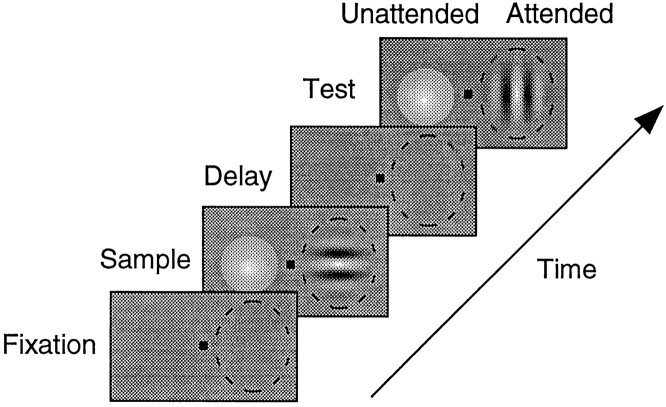
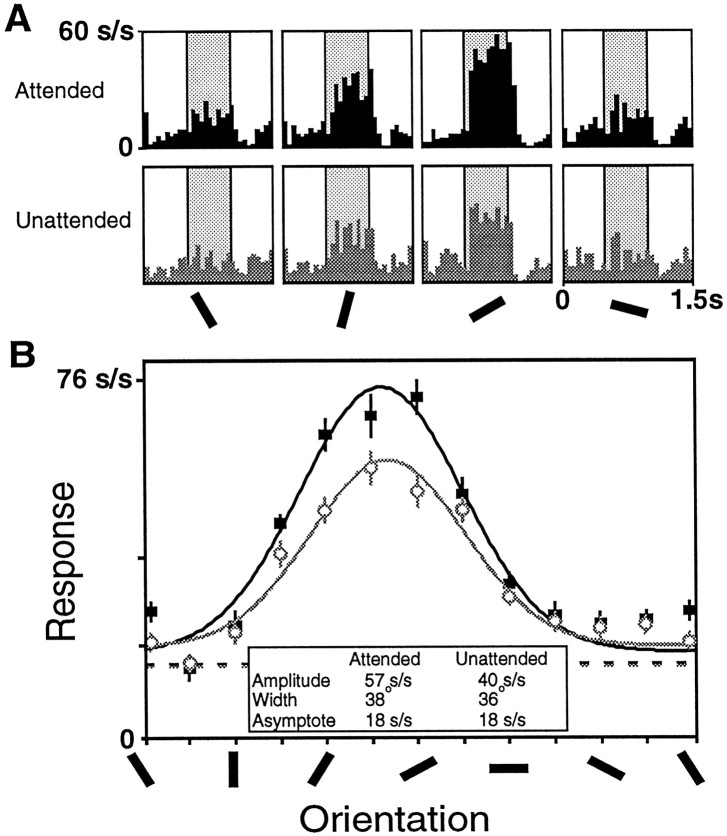
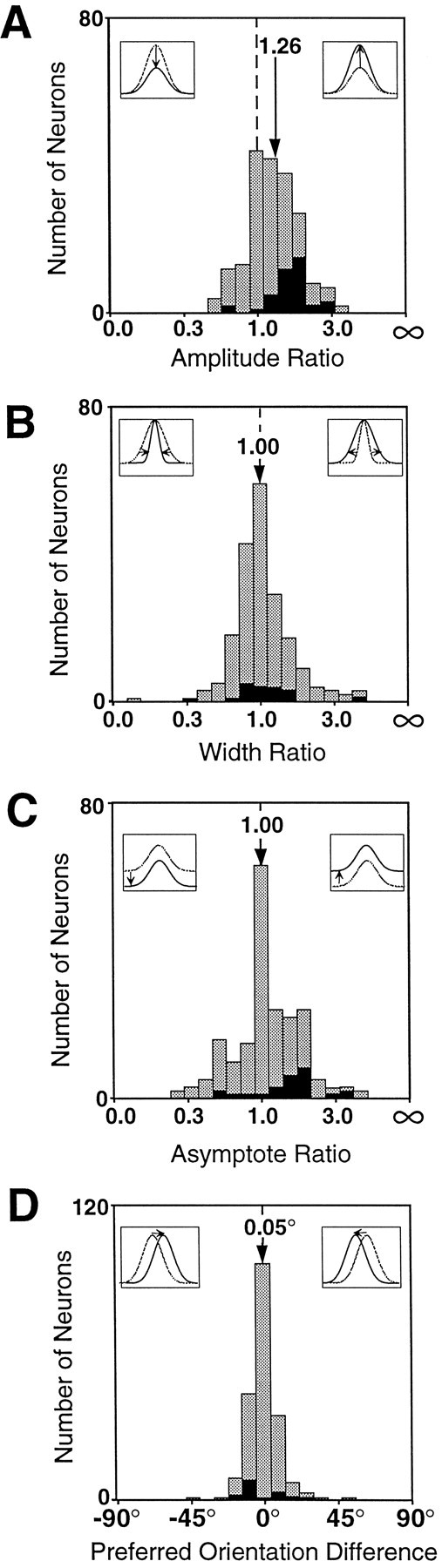
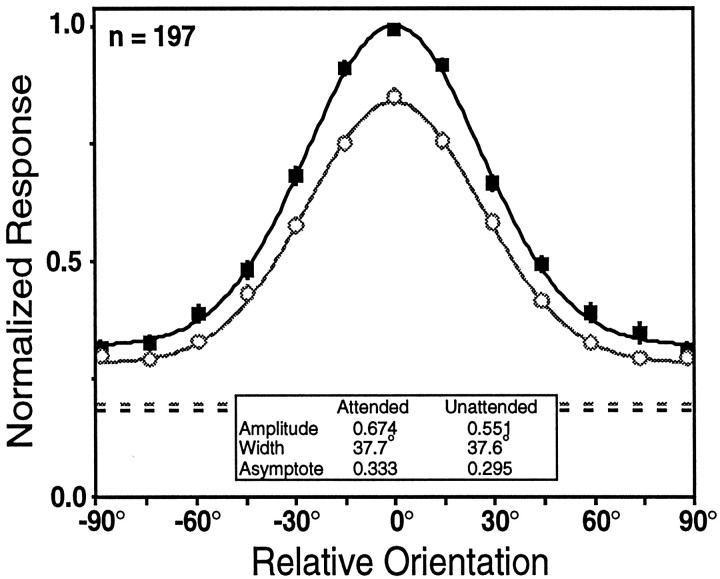
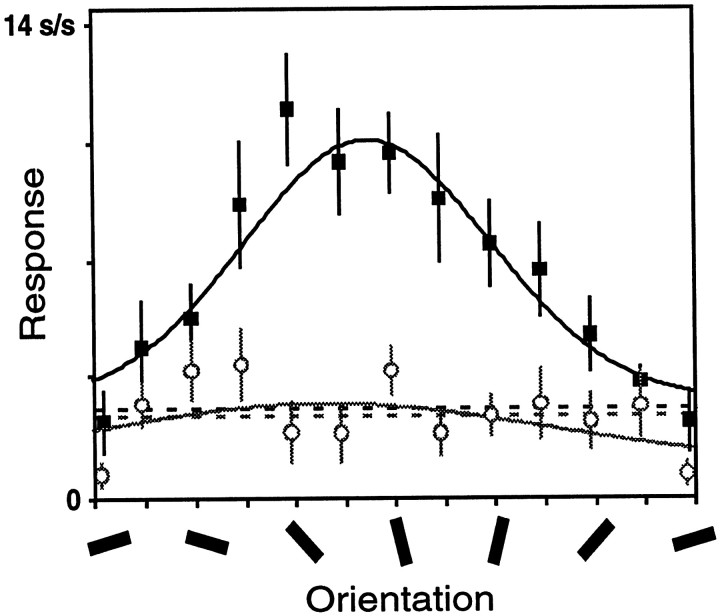
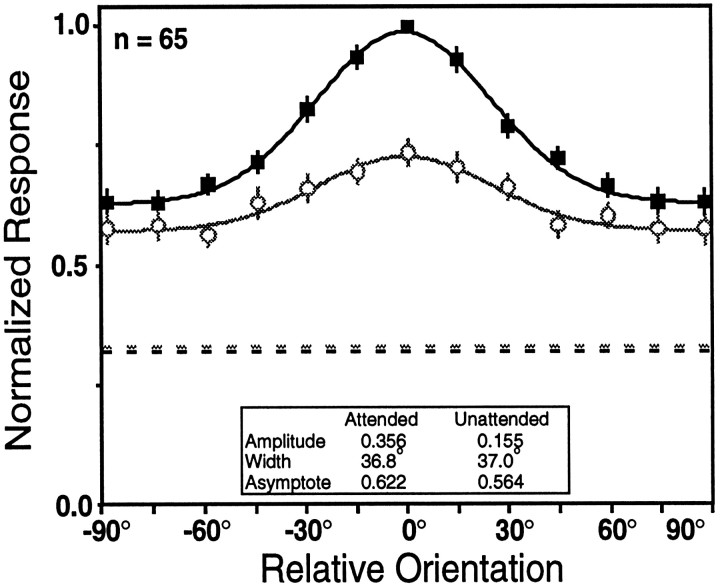
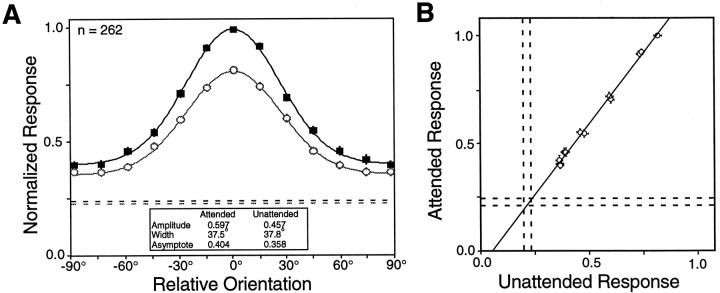
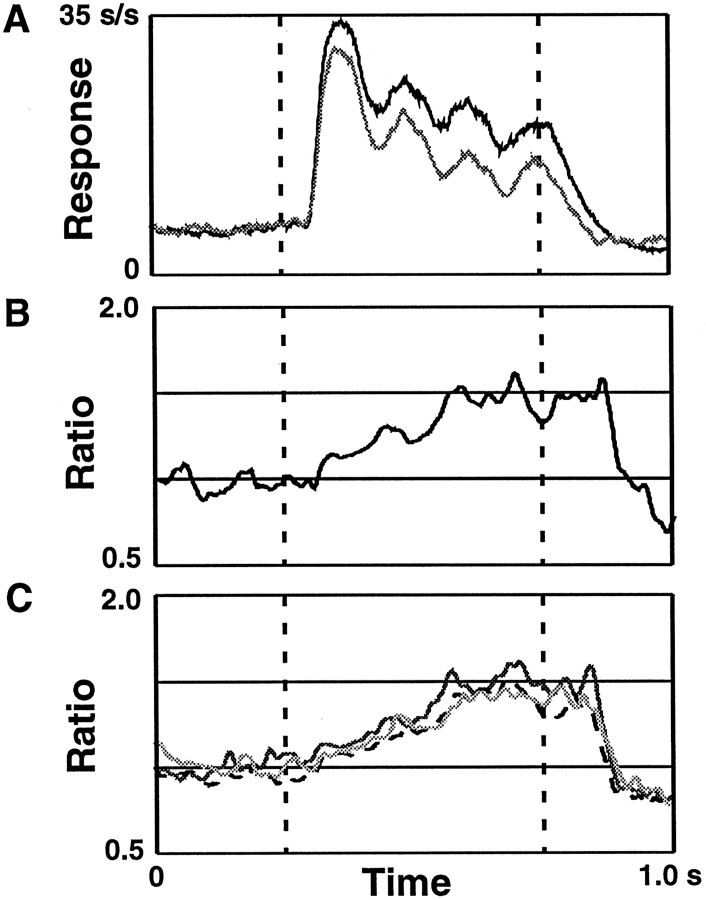
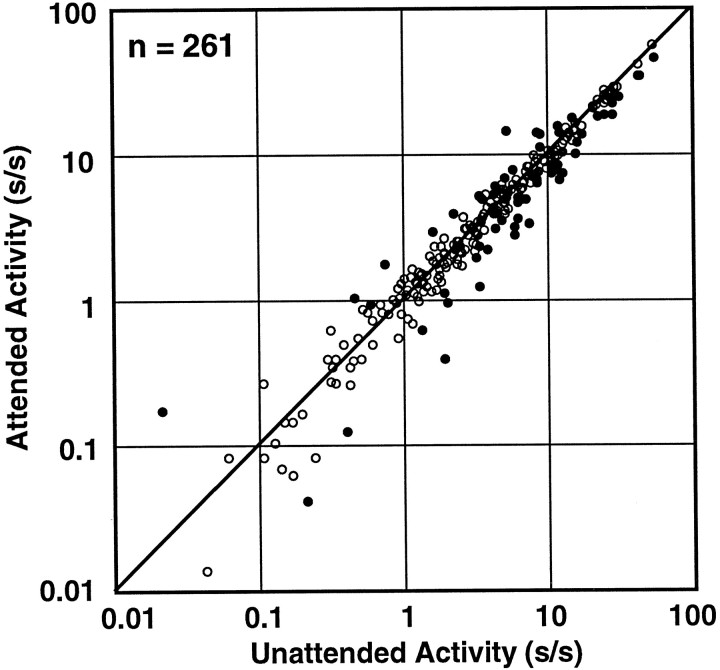
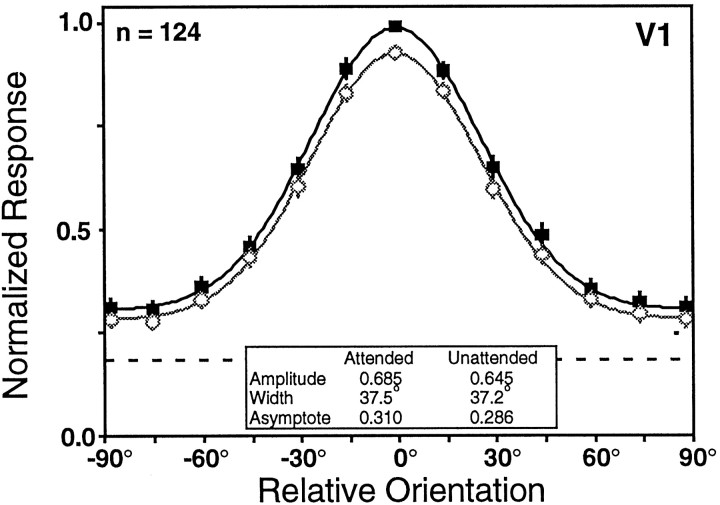
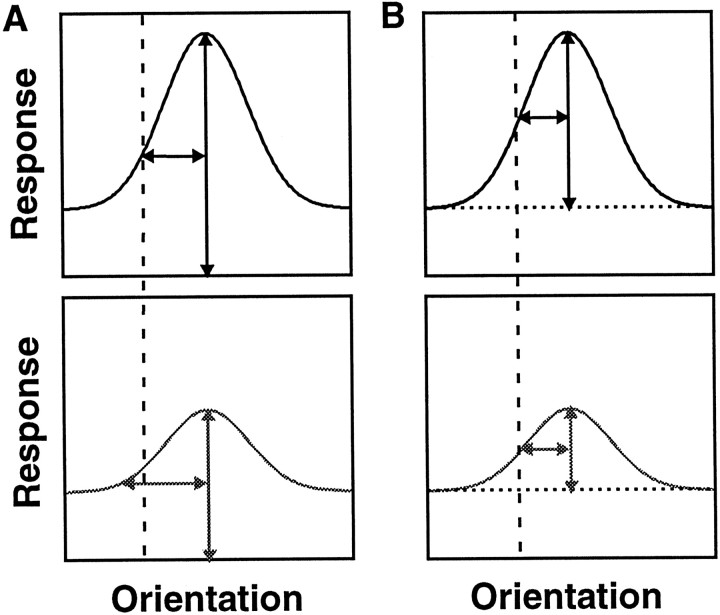
We examined how attention affected the orientation tuning of 262 isolated neurons in extrastriate area V4 and 135 neurons in area V1 of two rhesus monkeys. The animals were trained to perform a delayed match-to-sample task in which oriented stimuli were presented in the receptive field of the neuron being recorded. On some trials the animals were instructed to pay attention to those stimuli, and on other trials they were instructed to pay attention to other stimuli outside the receptive field. In this way, orientation-tuning curves could be constructed from neuronal responses collected in two behavioral states: one in which those stimuli were attended by the animal and one in which those stimuli were ignored by the animal. We fit Gaussians to the neuronal responses to twelve different orientations for each behavioral state. Although attention enhanced the responses of V4 neurons (median 26% increase) and V1 neurons (median 8% increase), selectivity, as measured by the width of its orientation-tuning curve, was not systematically altered by attention. The effects of attention were consistent with a multiplicative scaling of the driven response to all orientations. We also found that attention did not cause systematic changes in the undriven activity of the neurons.
Figures
References
-
- Adelson EH, Bergen JR. Spatiotemporal energy models for the perception of motion. J Opt Soc Am. 1985;2:284–299. - PubMed
-
- Albrecht DG, Geisler WS. Motion selectivity and the contrast-response function of simple cells in the visual cortex. Vis Neurosci. 1991;7:531–546. - PubMed
-
- Albrecht DG, Hamilton DB. Striate cortex of monkey and cat: contrast response function. J Neurophysiol. 1982;48:217–237. - PubMed
-
- Andersen RA, Essick GK, Siegel R. The encoding of spatial location by posterior parietal neurons. Science. 1985;230:456–458. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources