

Identification and characterization of novel superantigens from Streptococcus pyogenes
- PMID: 9874566
- PMCID: PMC1887688
- DOI: 10.1084/jem.189.1.89
Identification and characterization of novel superantigens from Streptococcus pyogenes
Abstract
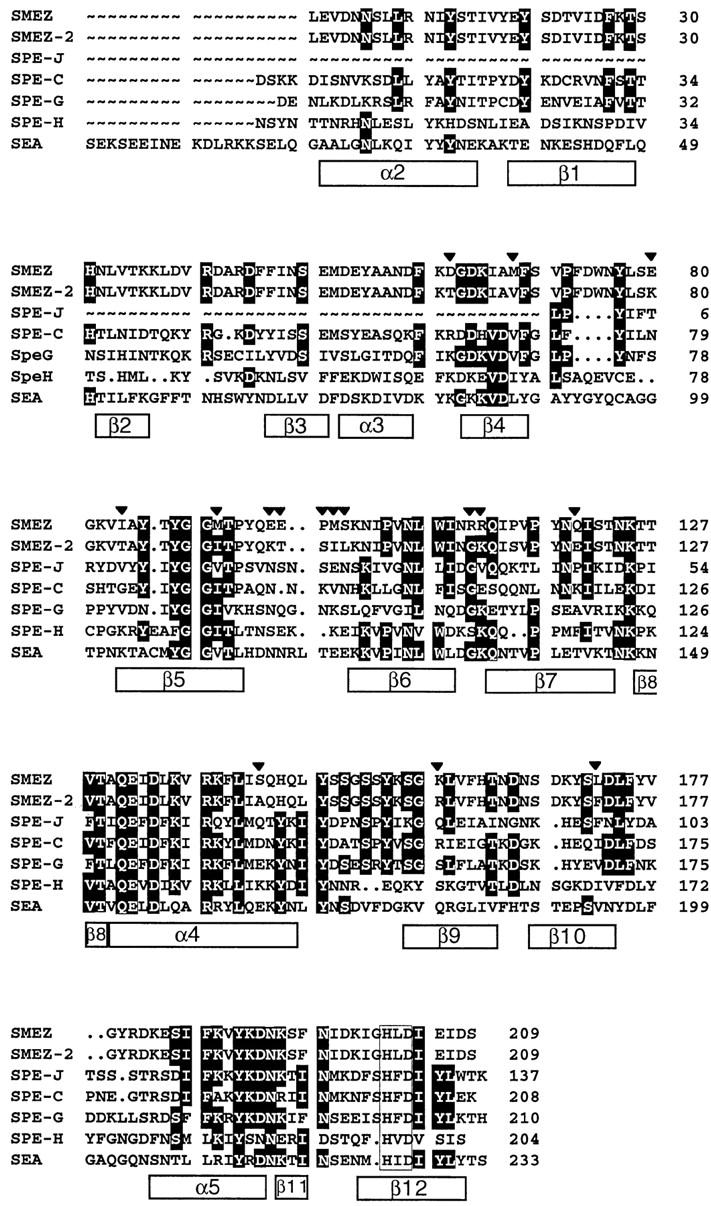
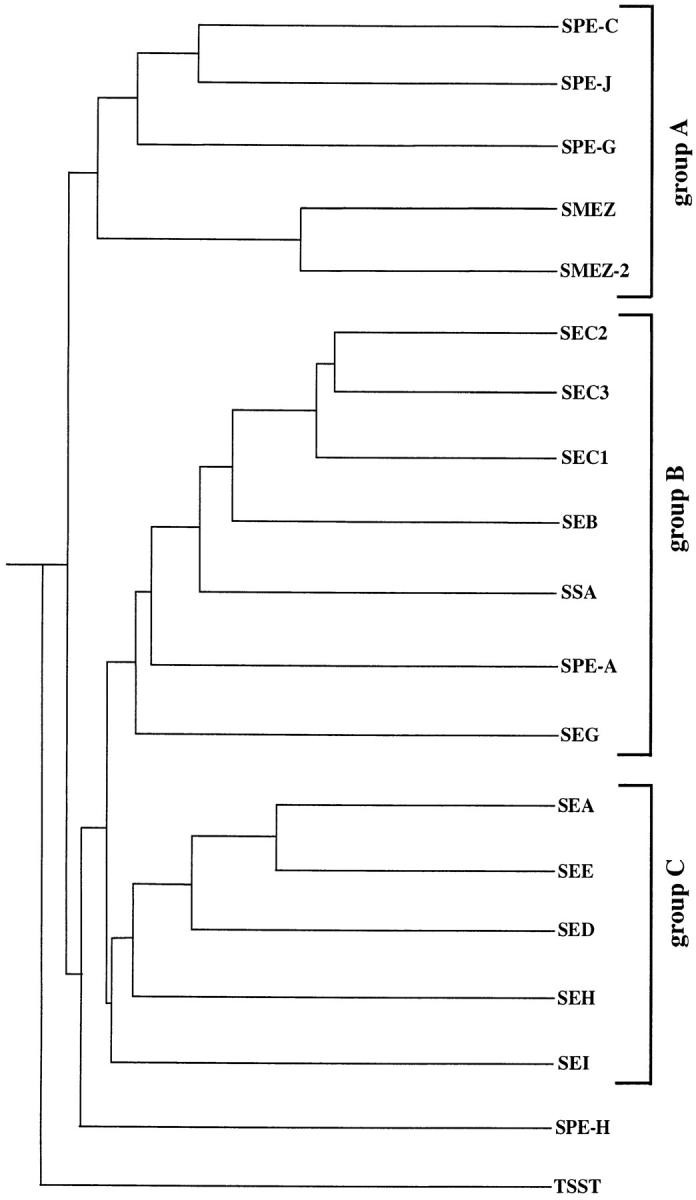
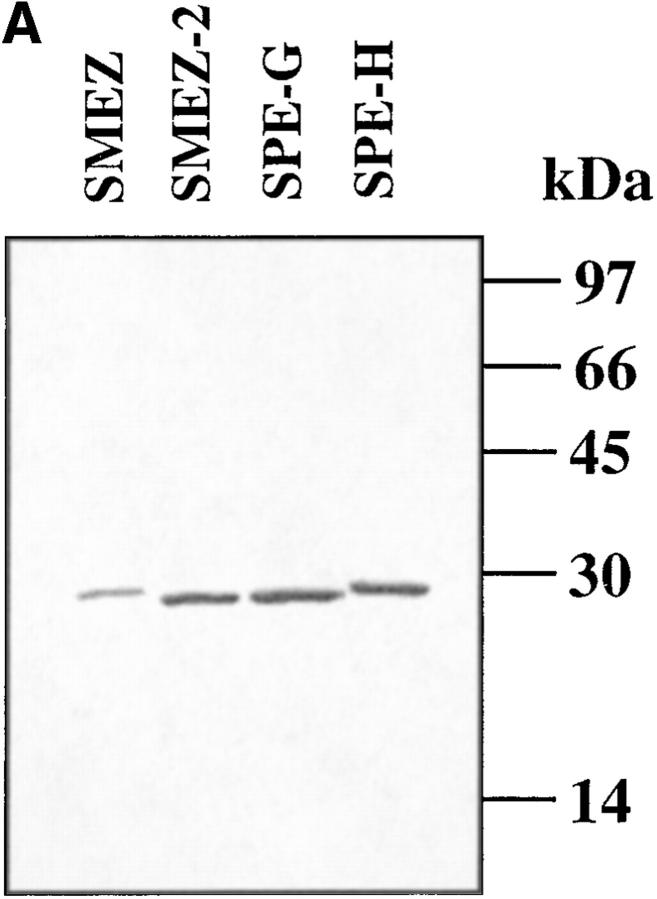
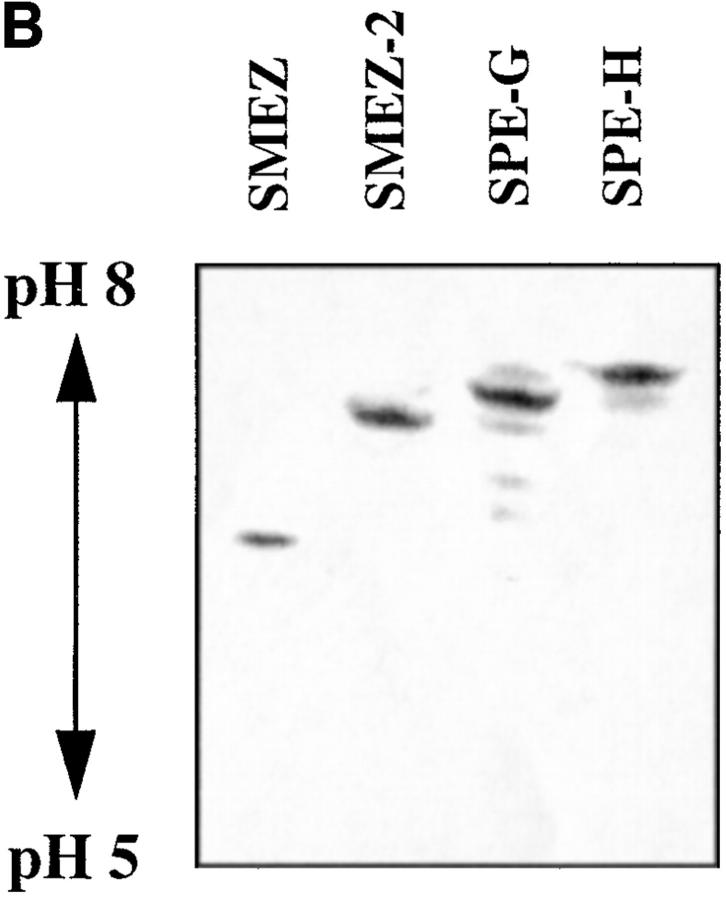
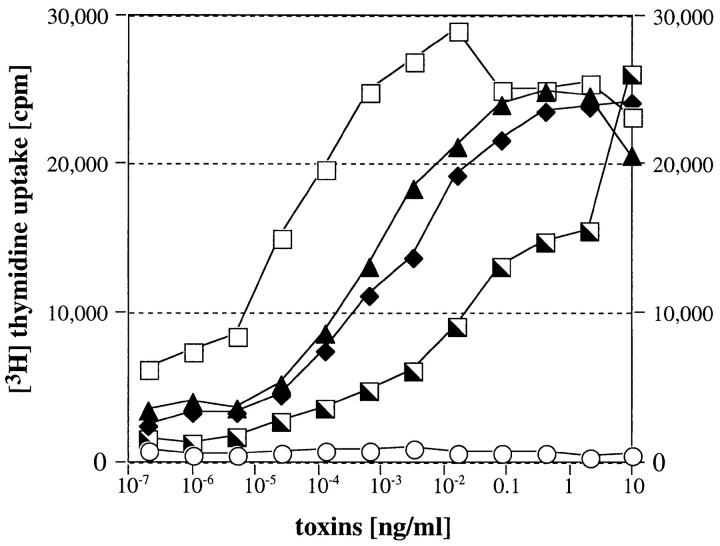
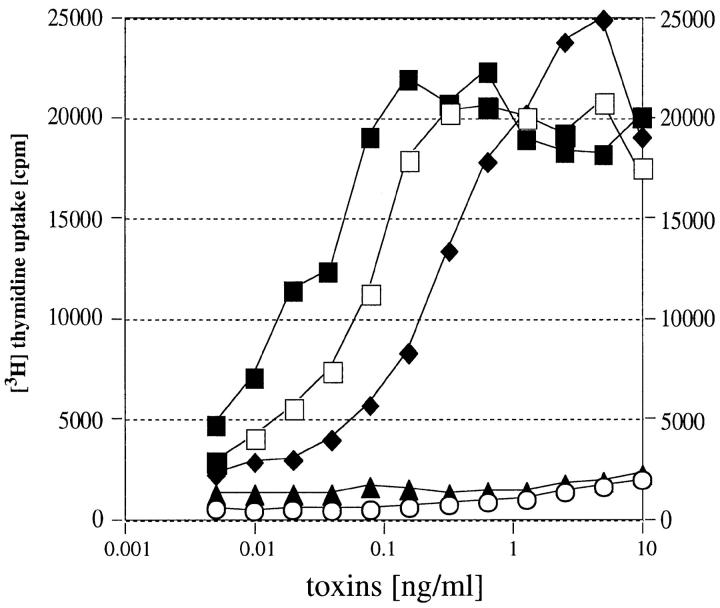
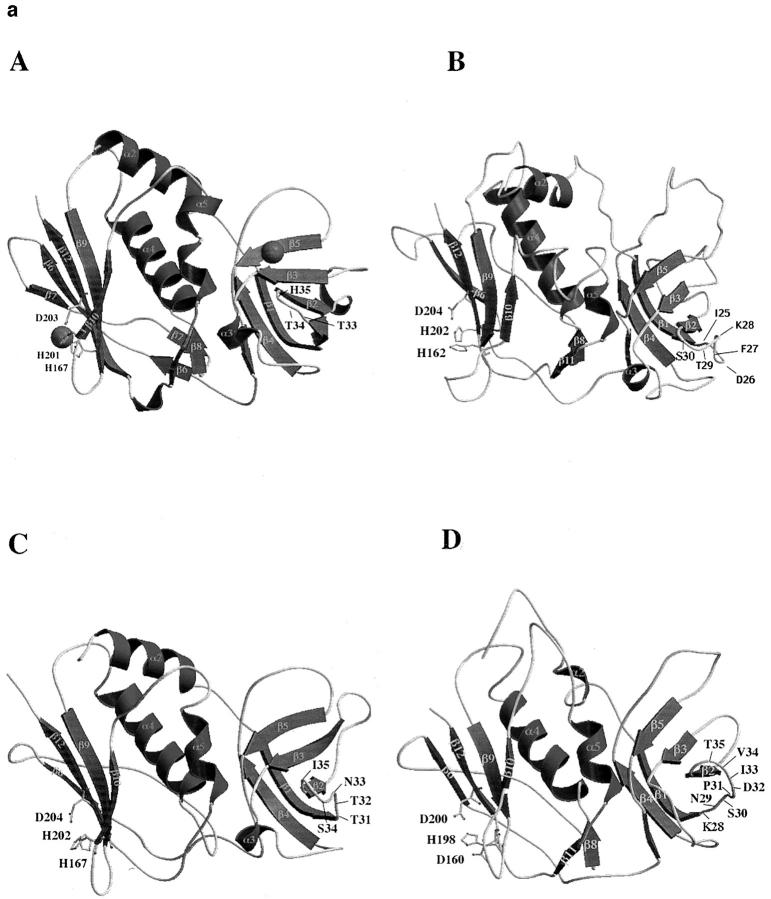
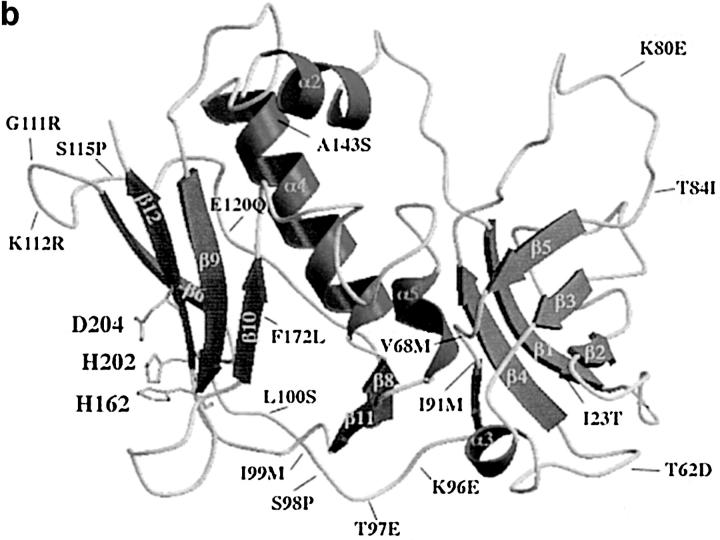
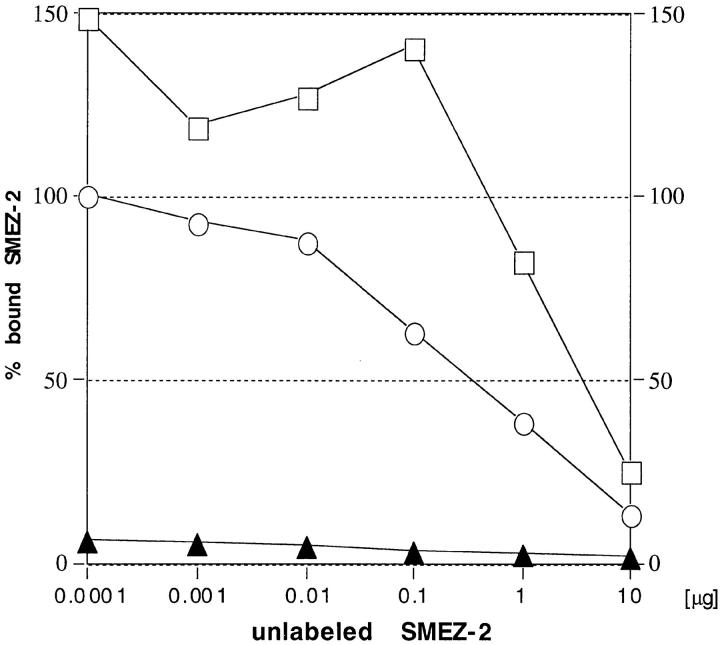
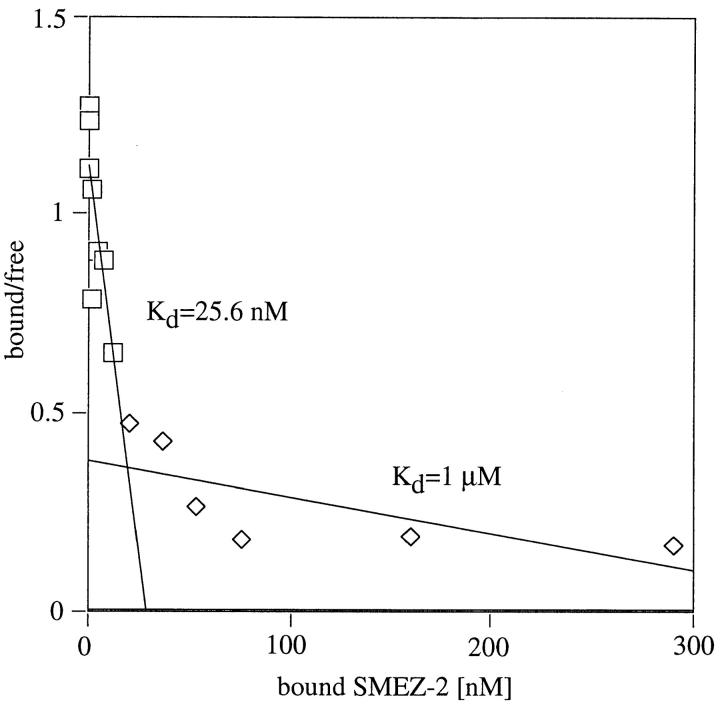
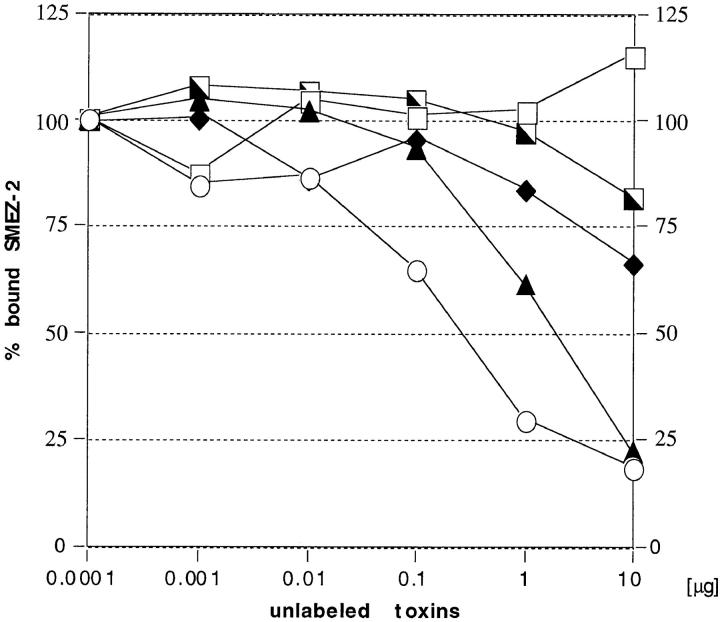
Three novel streptococcal superantigen genes (spe-g, spe-h, and spe-j) were identified from the Streptococcus pyogenes M1 genomic database at the University of Oklahoma. A fourth novel gene (smez-2) was isolated from the S. pyogenes strain 2035, based on sequence homology to the streptococcal mitogenic exotoxin z (smez) gene. SMEZ-2, SPE-G, and SPE-J are most closely related to SMEZ and streptococcal pyrogenic exotoxin (SPE)-C, whereas SPE-H is most similar to the staphylococcal toxins than to any other streptococcal toxin. Recombinant (r)SMEZ, rSMEZ-2, rSPE-G, and rSPE-H were mitogenic for human peripheral blood lymphocytes with half-maximal responses between 0.02 and 50 pg/ml (rSMEZ-2 and rSPE-H, respectively). SMEZ-2 is the most potent superantigen (SAg) discovered thus far. All toxins, except rSPE-G, were active on murine T cells, but with reduced potency. Binding to a human B-lymphoblastoid line was shown to be zinc dependent with high binding affinity of 15-65 nM. Evidence from modeled protein structures and competitive binding experiments suggest that high affinity binding of each toxin is to the major histocompatibility complex class II beta chain. Competition for binding between toxins was varied and revealed overlapping but discrete binding to subsets of class II molecules in the hierarchical order (SMEZ, SPE-C) > SMEZ-2 > SPE-H > SPE-G. The most common targets for the novel SAgs were human Vbeta2.1- and Vbeta4-expressing T cells. This might reflect a specific role for this subset of Vbetas in the immune defense of gram-positive bacteria.
Figures

References
-
- Marrack P, Kappler J. The staphylococcal enterotoxins and their relatives. Science. 1990;248:705–711. - PubMed
-
- Alouf, J.E., H. Knoell, and W. Koehler. 1991. The family of mitogenic, shock-inducing and superantigenic toxins from staphylococci and streptococci. In Sourcebook of Bacterial Protein Toxins. J.E. Alouf and J.H. Freer, editors. Academic Press, San Diego. 367–414.
-
- Betley MJ, Borst DW, Regassa LB. Staphylococcal enterotoxins, toxic shock syndrome toxin and streptococcal exotoxins: a comparative study of their molecular biology. Chem Immunol. 1992;55:1–35. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
