P2 receptor modulation of voltage-gated potassium currents in Brown adipocytes
- PMID: 9874693
- PMCID: PMC2222992
- DOI: 10.1085/jgp.113.1.125
P2 receptor modulation of voltage-gated potassium currents in Brown adipocytes
Abstract
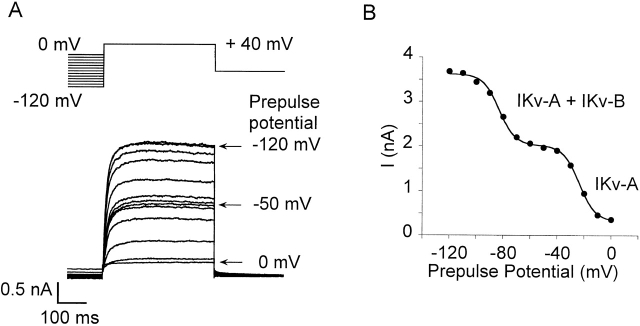
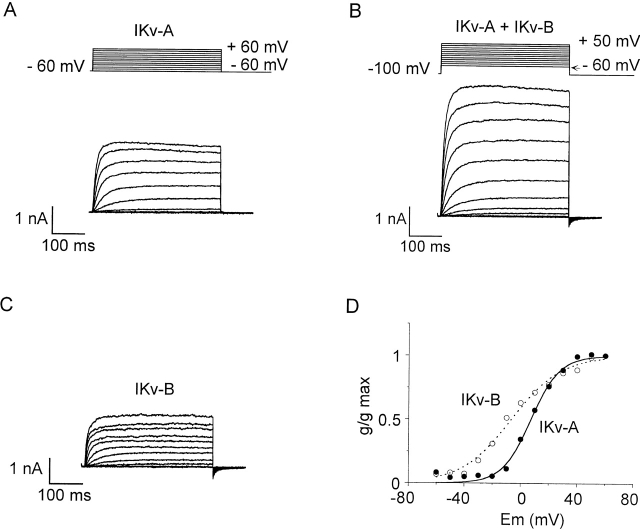
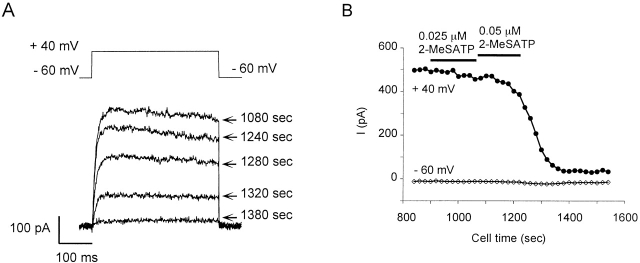
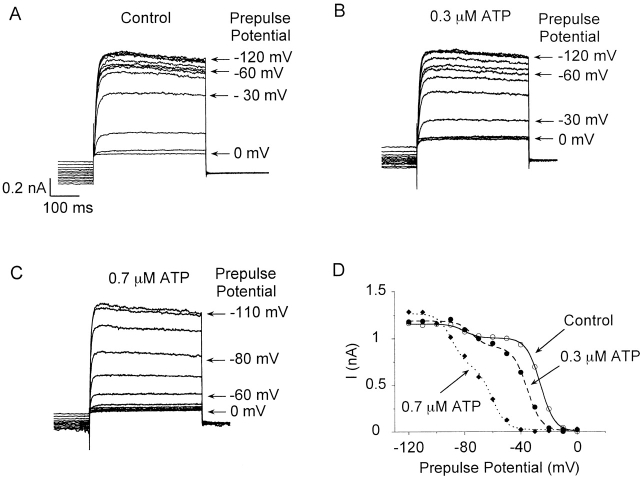
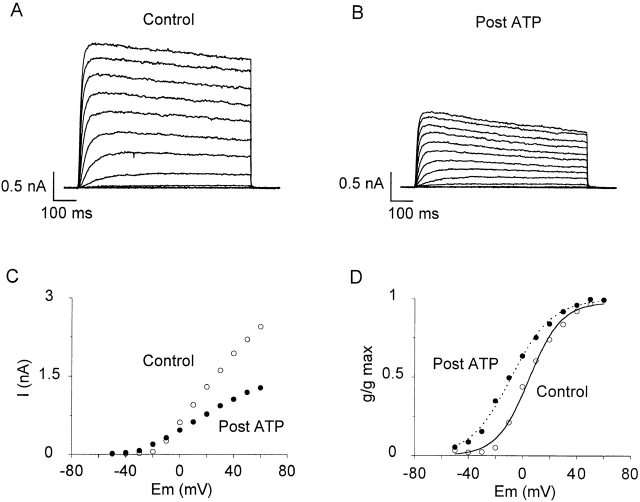
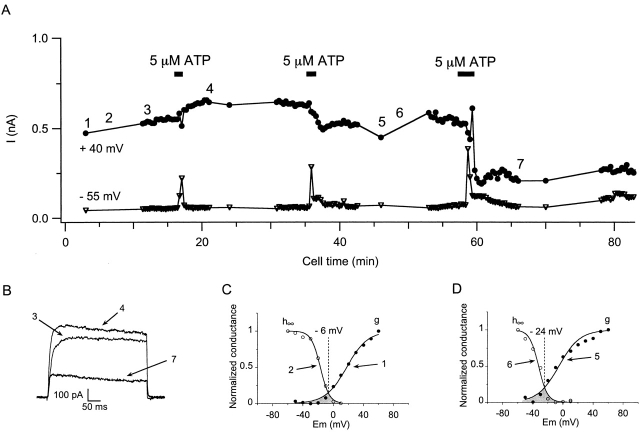
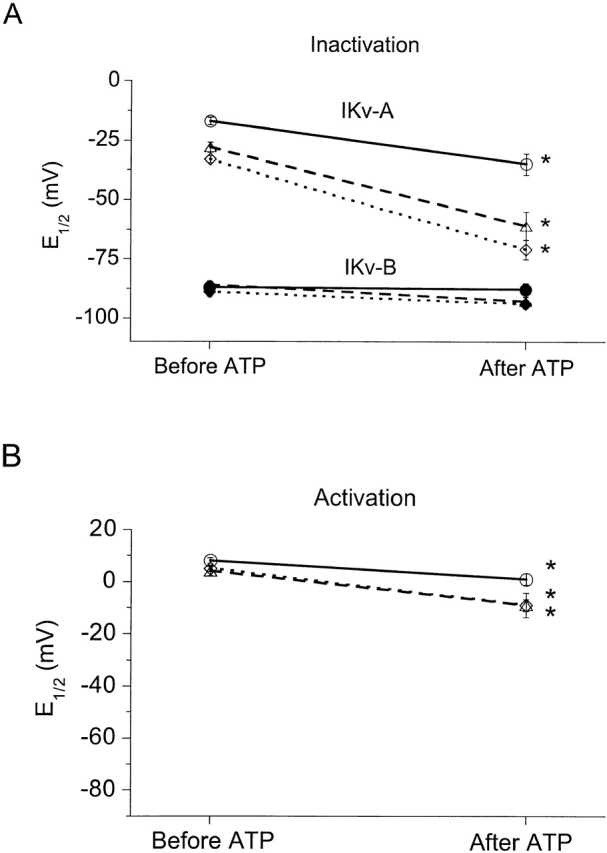
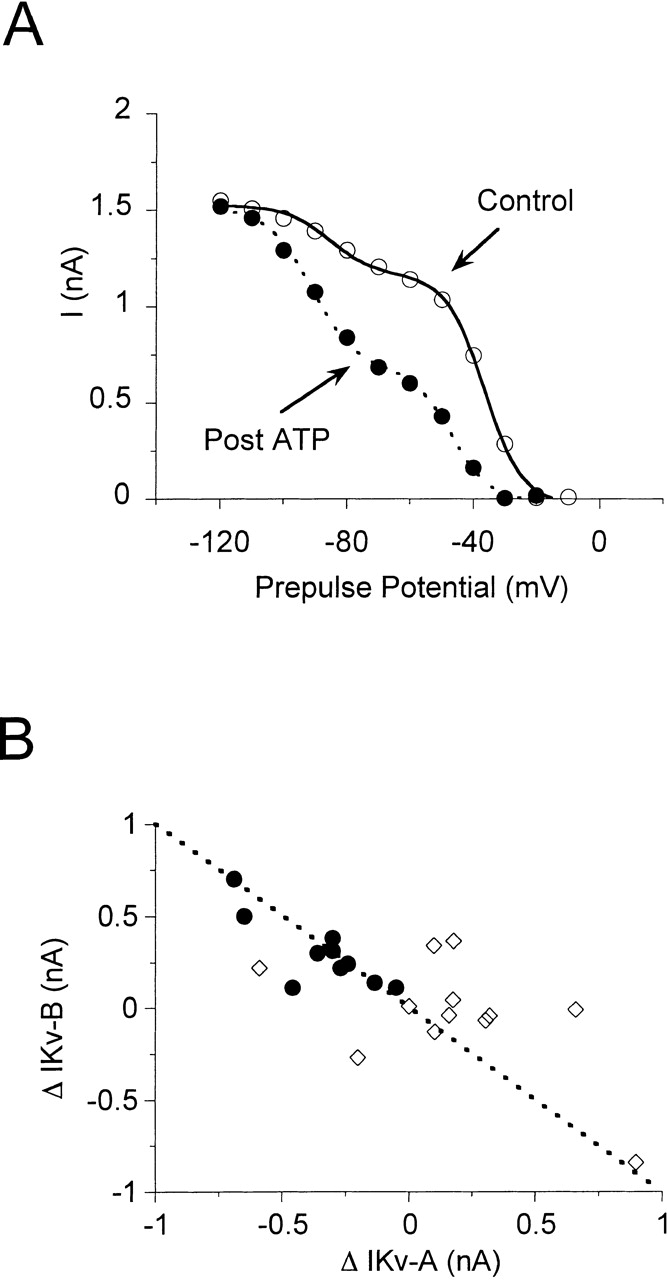
Using patch voltage-clamp techniques, we find there are two components to the voltage-gated potassium current (IKv) in rat brown adipocytes. The components differ in their gating and responses to purinergic stimulation, but not their pharmacology. IKv-A recovers from inactivation at physiological membrane potentials, while IKv-B inactivation recovers at more negative potentials. Both currents are >90% blocked by similar concentrations of quinine and tetraethylammonium, but not by beta-dendrotoxin, charybdotoxin, or apamin. The two current components are differentially modulated by extracellular ATP. ATP shifts the voltage dependence of IKv-A inactivation negative by 38 +/- 5 mV (n = 35, +/-SEM) and shifts activation by -14 +/- 2 mV in whole-cell experiments. ATP did not affect the steady state inactivation voltage dependence of IKv-B, but did apparently convert IKv-A into IKv-B. The pharmacology of the inactivation shift is consistent with mediation by a P2 purinergic receptor. Purinergic stimulation of perforated-patch clamped cells causes hyperpolarizing shifts in the window current of IKv-A by shifting inactivation -18 +/- 4 mV and activation -7 +/- 2 mV (n = 16). Since perforated-patch recordings will most closely resemble in vivo cell responses, this ATP-induced shift in the window current may facilitate IKv activation when the cell depolarizes. IKv activity is necessary for the proliferation and differentiation of brown adipocytes in culture (Pappone, P.A., and S.I. Ortiz-Miranda. 1993. Am. J. Physiol. 264:C1014-C1019) so purinergic modulation of IKv may be important in altering adipocyte growth and development.
Figures