

Alendronate mechanism of action: geranylgeraniol, an intermediate in the mevalonate pathway, prevents inhibition of osteoclast formation, bone resorption, and kinase activation in vitro
- PMID: 9874784
- PMCID: PMC15105
- DOI: 10.1073/pnas.96.1.133
Alendronate mechanism of action: geranylgeraniol, an intermediate in the mevalonate pathway, prevents inhibition of osteoclast formation, bone resorption, and kinase activation in vitro
Abstract
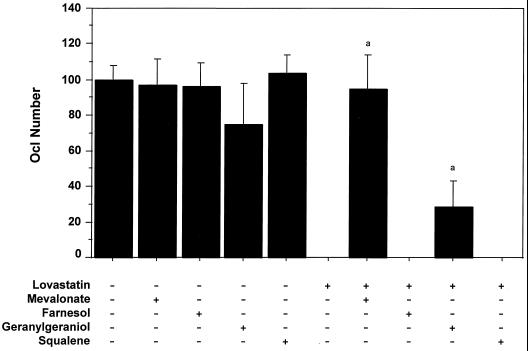
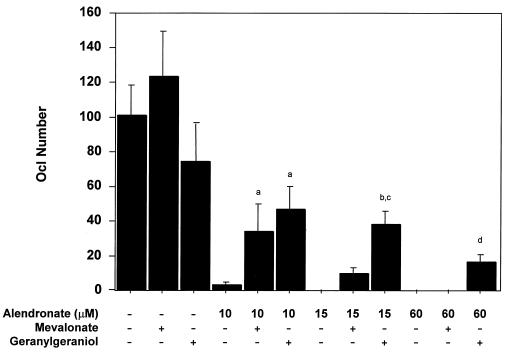
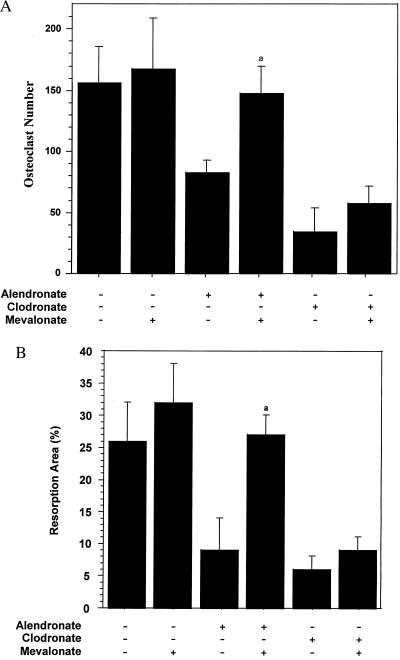
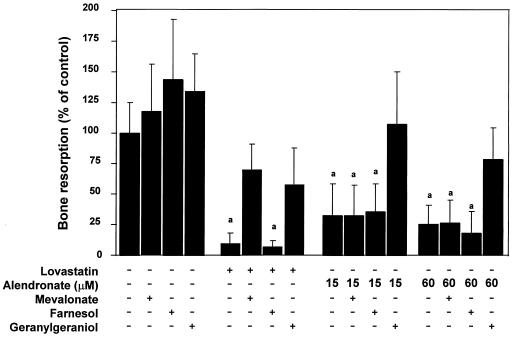
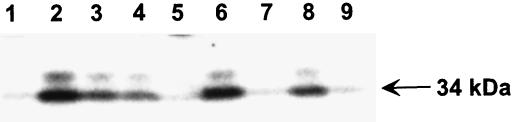
Nitrogen-containing bisphosphonates were shown to cause macrophage apoptosis by inhibiting enzymes in the biosynthetic pathway leading from mevalonate to cholesterol. This study suggests that, in osteoclasts, geranylgeranyl diphosphate, the substrate for prenylation of most GTP binding proteins, is likely to be the crucial intermediate affected by these bisphosphonates. We report that murine osteoclast formation in culture is inhibited by both lovastatin, an inhibitor of hydroxymethylglutaryl CoA reductase, and alendronate. Lovastatin effects are blocked fully by mevalonate and less effectively by geranylgeraniol whereas alendronate effects are blocked partially by mevalonate and more effectively by geranylgeraniol. Alendronate inhibition of bone resorption in mouse calvaria also is blocked by mevalonate whereas clodronate inhibition is not. Furthermore, rabbit osteoclast formation and activity also are inhibited by lovastatin and alendronate. The lovastatin effects are prevented by mevalonate or geranylgeraniol, and alendronate effects are prevented by geranylgeraniol. Farnesol and squalene are without effect. Signaling studies show that lovastatin and alendronate activate in purified osteoclasts a 34-kDa kinase. Lovastatin-mediated activation is blocked by mevalonate and geranylgeraniol whereas alendronate activation is blocked by geranylgeraniol. Together, these findings support the hypothesis that alendronate, acting directly on osteoclasts, inhibits a rate-limiting step in the cholesterol biosynthesis pathway, essential for osteoclast function. This inhibition is prevented by exogenous geranylgeraniol, probably required for prenylation of GTP binding proteins that control cytoskeletal reorganization, vesicular fusion, and apoptosis, processes involved in osteoclast activation and survival.
Figures
References
-
- Hughes D E, Wright K R, Uy H L, Sasaki A, Yoneda T, Roodman G D, Mundy G R, Boyce B F. J Bone Miner Res. 1995;10:1478–1487. - PubMed
-
- van Beek E, Hoekstra M, van der Ruit M, Lowik C, Papapoulos S. J Bone Miner Res. 1994;9:1875–1882. - PubMed
-
- Rogers M J, Xiong X, Brown R J, Watts D J, Russell R G G, Bayless A V, Ebetino F H. Mol Pharmacol. 1995;47:398–402. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
