

Association of Rab25 and Rab11a with the apical recycling system of polarized Madin-Darby canine kidney cells
- PMID: 9880326
- PMCID: PMC25153
- DOI: 10.1091/mbc.10.1.47
Association of Rab25 and Rab11a with the apical recycling system of polarized Madin-Darby canine kidney cells
Abstract
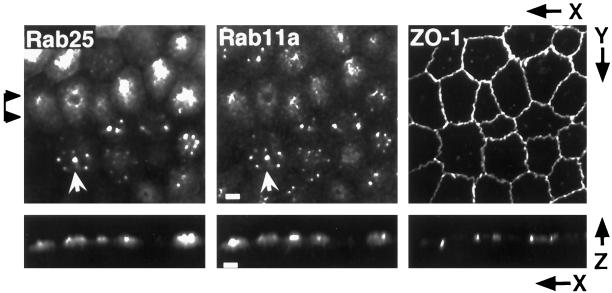
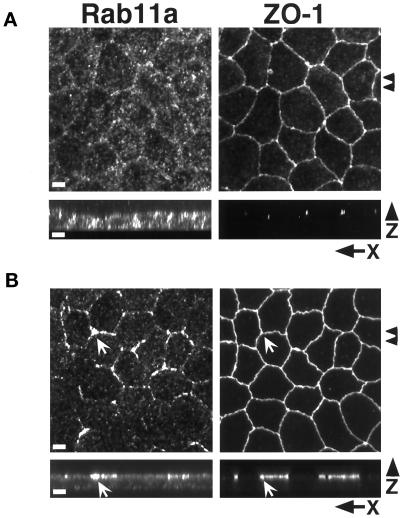
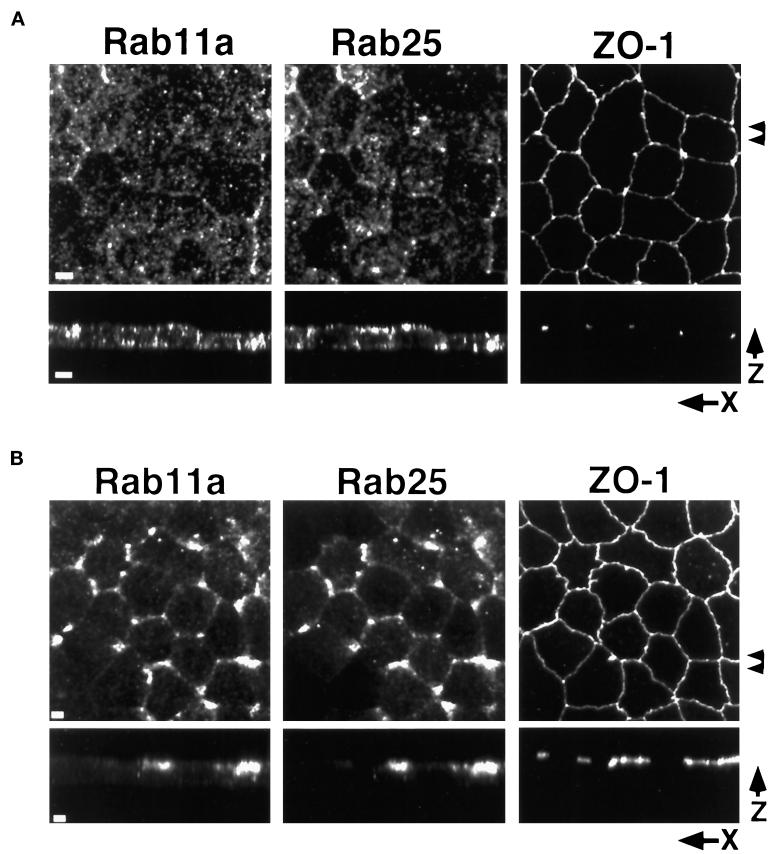
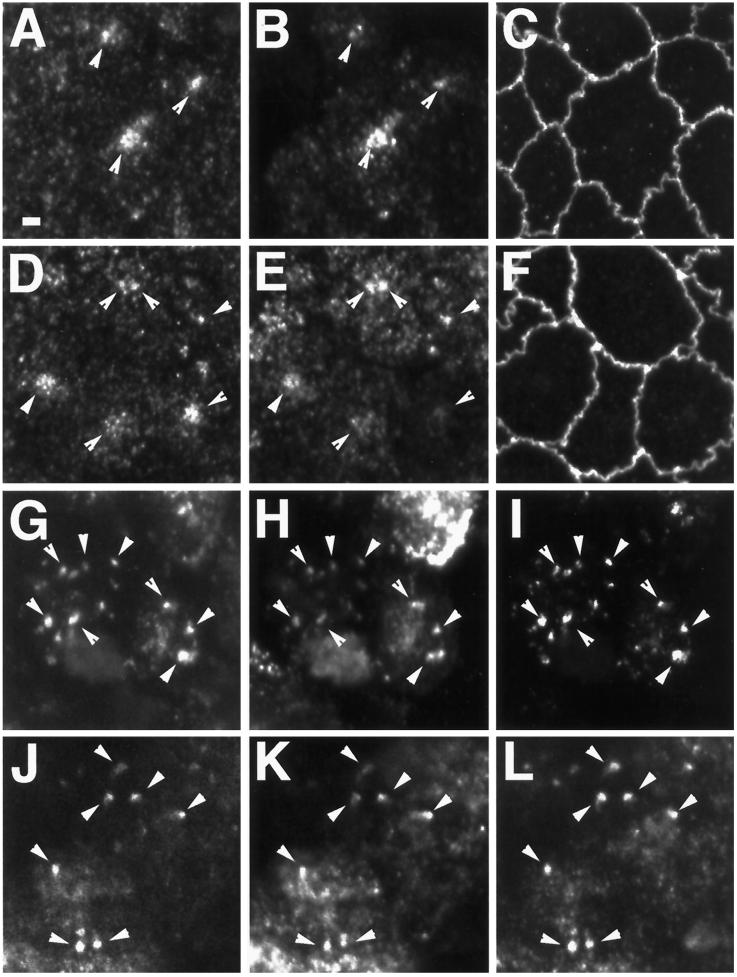
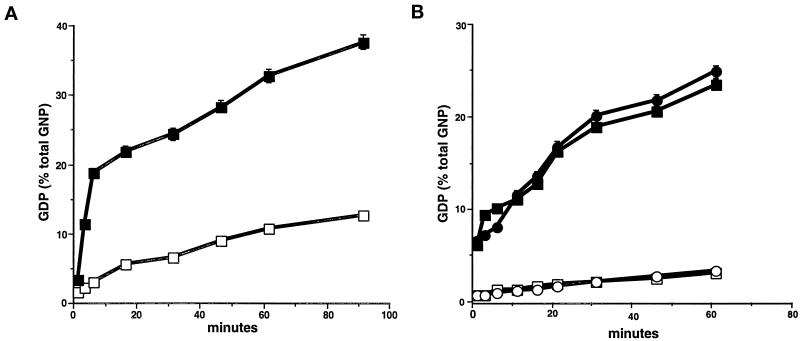
Recent evidence suggests that apical and basolateral endocytic pathways in epithelia converge in an apically located, pericentriolar endosomal compartment termed the apical recycling endosome. In this compartment, apically and basolaterally internalized membrane constituents are thought to be sorted for recycling back to their site of origin or for transcytosis to the opposite plasma membrane domain. We report here that in the epithelial cell line Madin-Darby Canine Kidney (MDCK), antibodies to Rab11a label an apical pericentriolar endosomal compartment that is dependent on intact microtubules for its integrity. Furthermore, this compartment is accessible to a membrane-bound marker (dimeric immunoglobulin A [IgA]) internalized from either the apical or basolateral pole, functionally defining it as the apical recycling endosome. We have also examined the role of a closely related epithelial-specific Rab, Rab25, in the regulation of membrane recycling and transcytosis in MDCK cells. When cDNA encoding Rab25 was transfected into MDCK cells, the protein colocalized with Rab11a in subapical vesicles. Rab25 transfection also altered the distribution of Rab11a, causing the coalescence of immunoreactivity into multiple denser vesicular structures not associated with the centrosome. Nevertheless, nocodazole still dispersed these vesicles, and dimeric IgA internalized from either the apical or basolateral membrane was detected in endosomes labeled with antibodies to both Rab11a and Rab25. Overexpression of Rab25 decreased the rate of IgA transcytosis and of apical, but not basolateral, recycling of internalized ligand. Conversely, expression of the dominant-negative Rab25T26N did not alter either apical recycling or transcytosis. These results indicate that both Rab11a and Rab25 associate with the apical recycling system of epithelial cells and suggest that Rab25 may selectively regulate the apical recycling and/or transcytotic pathways.
Figures



References
-
- Adari H, Lowy DR, Willumsen BM, Der CJ, McCormick F. Guanosine triphosphatase activating protein (GAP) interacts with the p21 ras effector binding domain. Science. 1988;240:518–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
