

Dynamics of cytokinins in apical shoot meristems of a day-neutral tobacco during floral transition and flower formation
- PMID: 9880352
- PMCID: PMC32210
- DOI: 10.1104/pp.119.1.111
Dynamics of cytokinins in apical shoot meristems of a day-neutral tobacco during floral transition and flower formation
Abstract
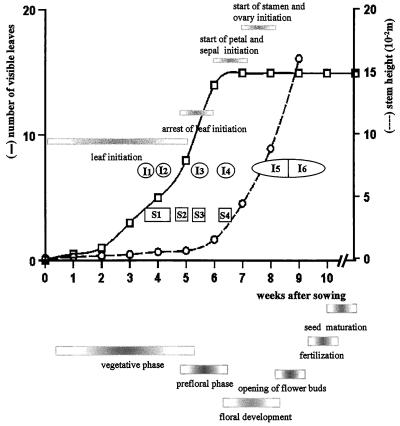
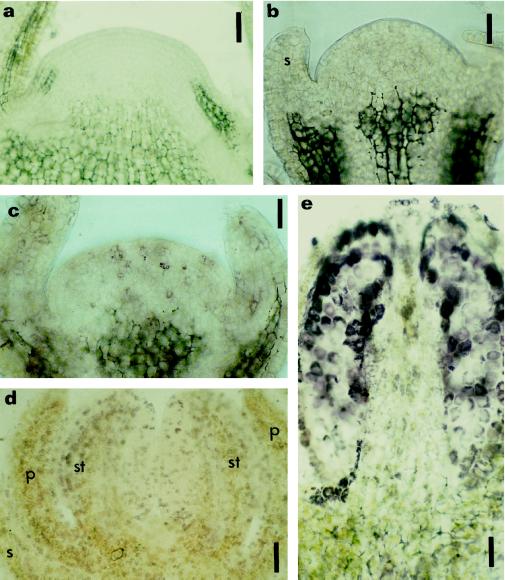
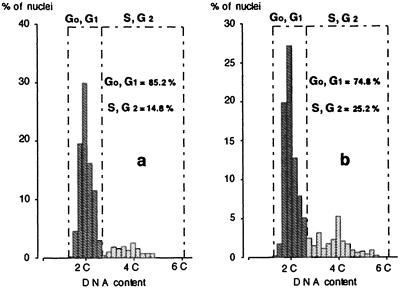
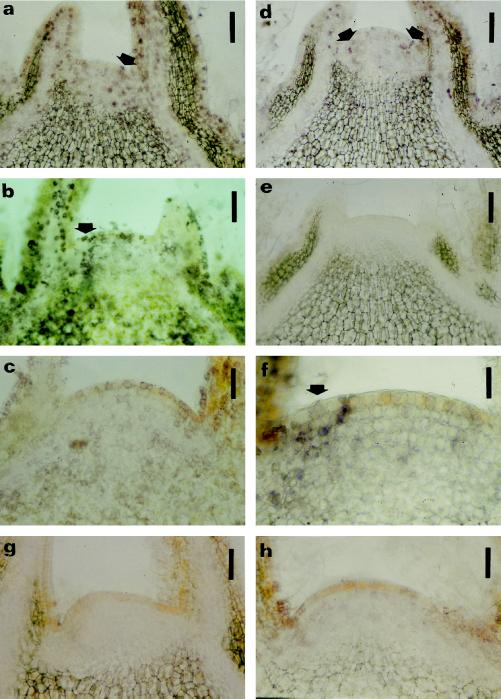
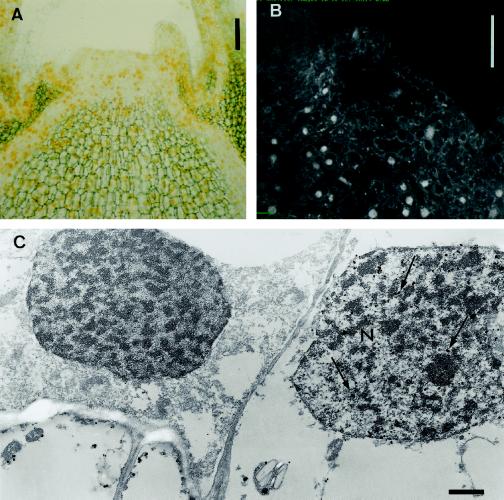
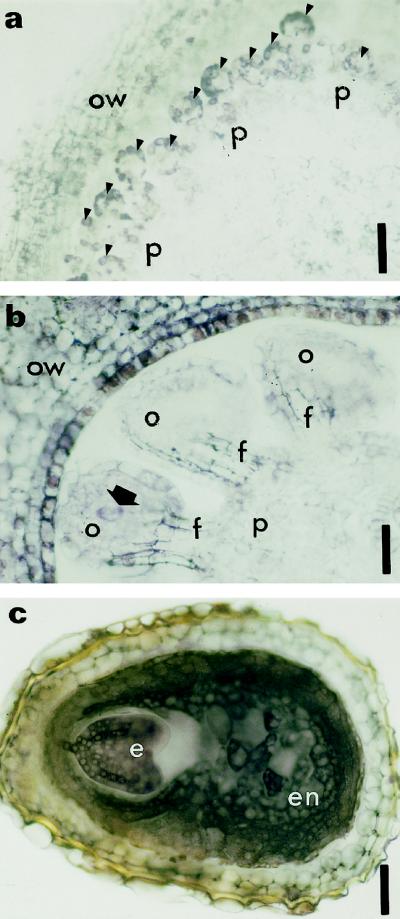
This study considered cytokinin distribution in tobacco (Nicotiana tabacum L.) shoot apices in distinct phases of development using immunocytochemistry and quantitative tandem mass spectrometry. In contrast to vegetative apices and flower buds, we detected no free cytokinin bases (zeatin, dihydrozeatin, or isopentenyladenine) in prefloral transition apices. We also observed a 3-fold decrease in the content of cytokinin ribosides (zeatin riboside, dihydrozeatin riboside, and isopentenyladenosine) during this transition phase. The group concluded that organ formation (e.g. leaves and flowers) is characterized by enhanced cytokinin content, in contrast to the very low endogenous cytokinin levels found in prefloral transition apices, which showed no organogenesis. The immunocytochemical analyses revealed a differing intracellular localization of the cytokinin bases. Dihydrozeatin and isopentenyladenine were mainly cytoplasmic and perinuclear, whereas zeatin showed a clear-cut nuclear labeling. To our knowledge, this is the first time that this phenomenon has been reported. Cytokinins do not seem to act as positive effectors in the prefloral transition phase in tobacco shoot apices. Furthermore, the differences in distribution at the cellular level may be indicative of a specific physiological role of zeatin in nuclear processes.
Figures
References
-
- Bernier G. The control of floral evocation and morphogenesis. Annu Rev Plant Physiol Plant Mol Biol. 1988;39:175–219.
-
- Besnard-Wibaut C. Effectiveness of gibberellins and 6-benzyladenine of flowering of Arabidopsis thaliana. Physiol Plant. 1981;53:205–212.
-
- Beveridge CA, Murfet IC, Kerhoas L, Sotta B, Miginiac E, Rameau C. The shoot controls zeatin riboside export from pea roots: evidence from the branching mutant rms4. Plant J. 1997;11:339–345.
-
- Biermann BJ, Morehead AT, Tate ST, Price JF, Randhall SK, Crowell DN. Novel isoprenylated proteins identified by an expression library screen. J Biol Chem. 1994;269:25251–25254. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
