

Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore-deficient aurea and yellow-green-2 mutants of tomato
- PMID: 9880355
- PMCID: PMC32213
- DOI: 10.1104/pp.119.1.143
Feedback inhibition of chlorophyll synthesis in the phytochrome chromophore-deficient aurea and yellow-green-2 mutants of tomato
Abstract
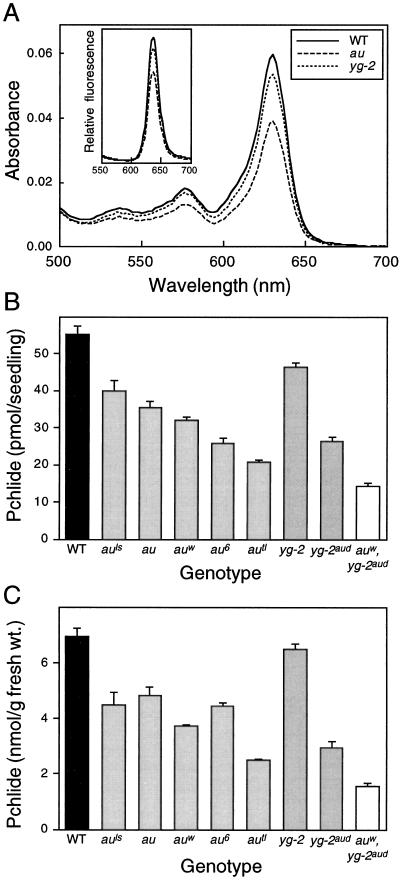
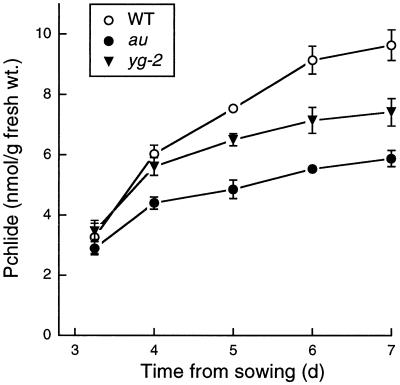
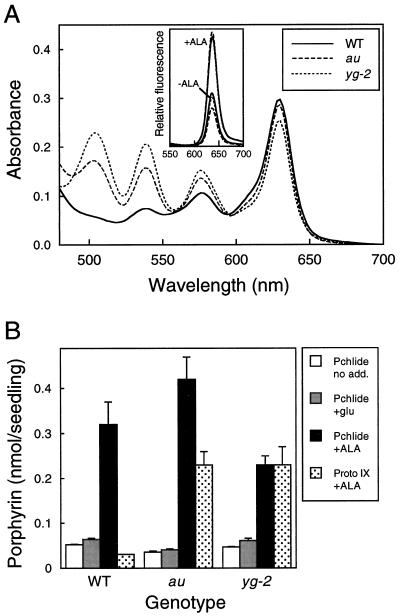
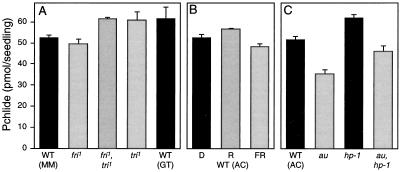
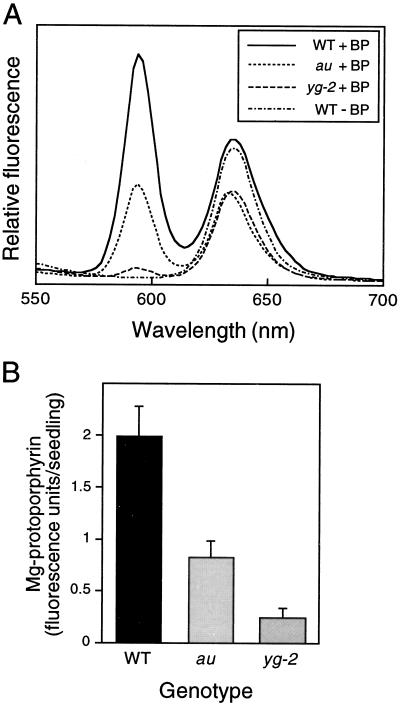
The aurea (au) and yellow-green-2 (yg-2) mutants of tomato (Solanum lycopersicum L.) are unable to synthesize the linear tetrapyrrole chromophore of phytochrome, resulting in plants with a yellow-green phenotype. To understand the basis of this phenotype, we investigated the consequences of the au and yg-2 mutations on tetrapyrrole metabolism. Dark-grown seedlings of both mutants have reduced levels of protochlorophyllide (Pchlide) due to an inhibition of Pchlide synthesis. Feeding experiments with the tetrapyrrole precursor 5-aminolevulinic acid (ALA) demonstrate that the pathway between ALA and Pchlide is intact in au and yg-2 and suggest that the reduction in Pchlide is a result of the inhibition of ALA synthesis. This inhibition was independent of any deficiency in seed phytochrome, and experiments using an iron chelator to block heme synthesis demonstrated that both mutations inhibited the degradation of the physiologically active heme pool, suggesting that the reduction in Pchlide synthesis is a consequence of feedback inhibition by heme. We discuss the significance of these results in understanding the chlorophyll-deficient phenotype of the au and yg-2 mutants.
Figures

References
-
- Adamse P, Peters JL, Jaspers PAPM, van Tuinen A, Koornneef M, Kendrick RE. Photocontrol of anthocyanin synthesis in tomato seedlings: a genetic approach. Photochem Photobiol. 1989;50:107–111.
-
- Batschauer A, Gilmartin PM, Nagy F, Schäfer E. The molecular biology of photoregulated genes. In: Kendrick RE, Kronenberg GHM, editors. Photomorphogenesis in Plants, Ed 2. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1994. pp. 559–600.
-
- Beale SI, Weinstein JD. Biochemistry and regulation of photosynthetic pigment formation in plants and algae. In: Jordan PM, editor. Biosynthesis of Tetrapyrroles. Amsterdam, The Netherlands: Elsevier; 1991. pp. 155–235.
-
- Becker TW, Foyer C, Caboche M. Light-regulated expression of the nitrate-reductase and nitrite-reductase genes in tomato and in the phytochrome-deficient aurea mutant of tomato. Planta. 1992;188:39–47. - PubMed
LinkOut - more resources
Full Text Sources
