

An in vitro system from maize seedlings for tryptophan-independent indole-3-acetic acid biosynthesis
- PMID: 9880358
- PMCID: PMC32217
- DOI: 10.1104/pp.119.1.173
An in vitro system from maize seedlings for tryptophan-independent indole-3-acetic acid biosynthesis
Abstract
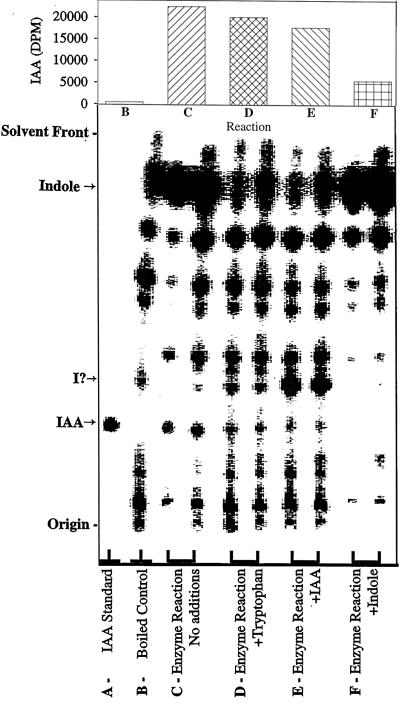
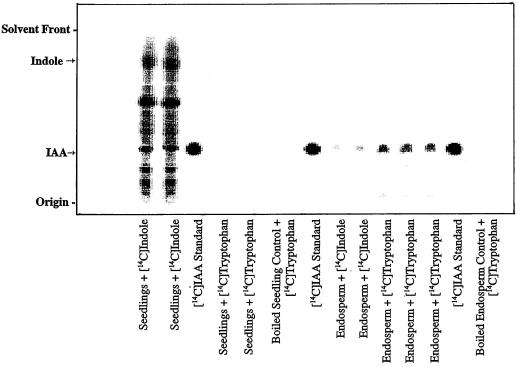
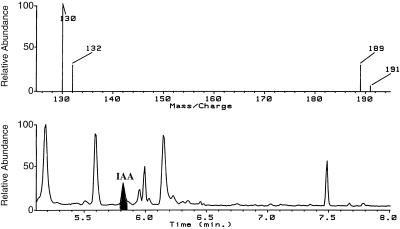
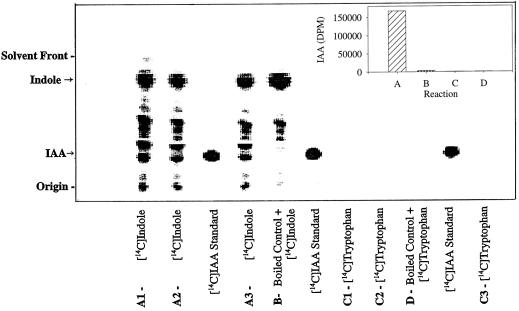
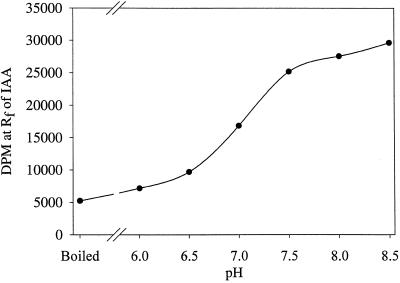
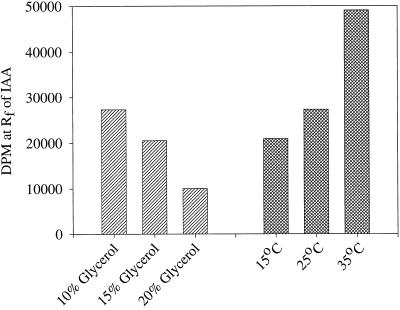
The enzymatic synthesis of indole-3-acetic acid (IAA) from indole by an in vitro preparation from maize (Zea mays L.) that does not use tryptophan (Trp) as an intermediate is described. Light-grown seedlings of normal maize and the maize mutant orange pericarp were shown to contain the necessary enzymes to convert [14C]indole to IAA. The reaction was not inhibited by unlabeled Trp and neither [14C]Trp nor [14C]serine substituted for [14C]indole in this in vitro system. The reaction had a pH optimum greater than 8.0, required a reducing environment, and had an oxidation potential near that of ascorbate. The results obtained with this in vitro enzyme preparation provide strong, additional evidence for the presence of a Trp-independent IAA biosynthesis pathway in plants.
Figures
References
-
- Bandurski RS, Cohen JD, Slovin JP, Reinecke DM. Auxin biosynthesis and metabolism. In: Davies PJ, editor. Plant Hormones: Physiology, Biochemistry and Molecular Biology. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1995. pp. 35–57.
-
- Bartel B. Auxin biosynthesis. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:51–66. - PubMed
-
- Chauhan YS, Rathore VS, Garg GK, Bhargava A. Detection of an indole oxidizing system in maize leaves. Biochem Biophys Res Commun. 1978;83:1237–1245. - PubMed
-
- Cohen JD, Bialek K. The biosynthesis of indole-3-acetic acid in higher plants. In: Crozier A, Hillman JR, editors. The Biosynthesis and Metabolism of Plant Hormones. Society for Experimental Biology Seminar 23. Cambridge, UK: Cambridge University Press; 1984. pp. 165–181.
LinkOut - more resources
Full Text Sources
