

Regulation of S-like ribonuclease levels in Arabidopsis. Antisense inhibition of RNS1 or RNS2 elevates anthocyanin accumulation
- PMID: 9880376
- PMCID: PMC32237
- DOI: 10.1104/pp.119.1.331
Regulation of S-like ribonuclease levels in Arabidopsis. Antisense inhibition of RNS1 or RNS2 elevates anthocyanin accumulation
Abstract
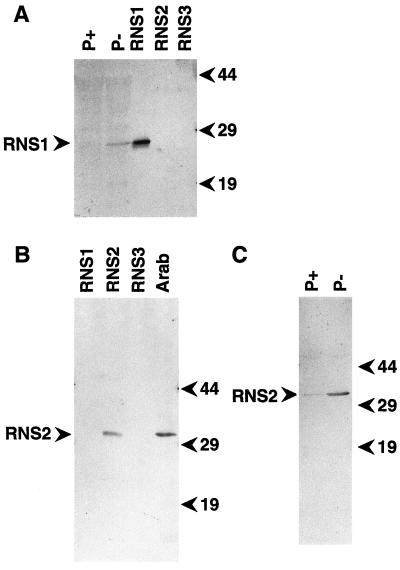
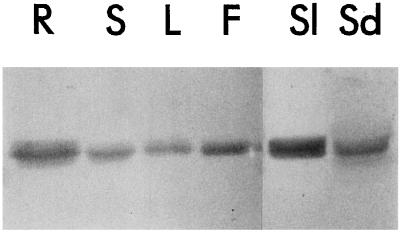
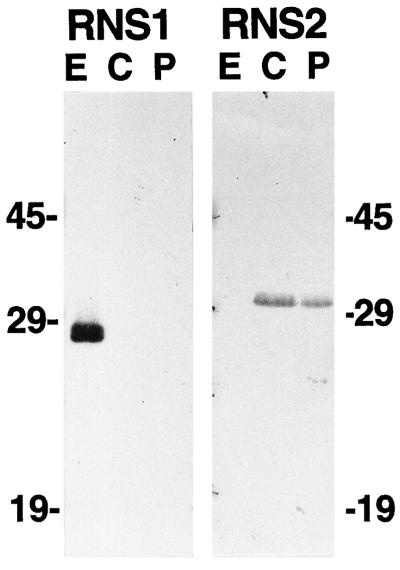
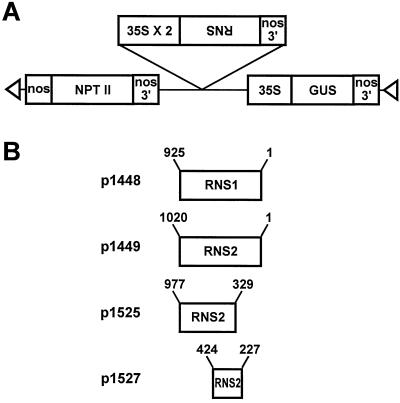
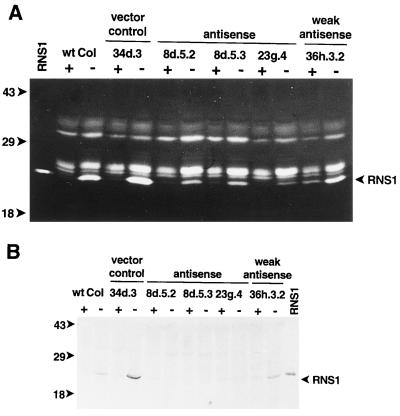
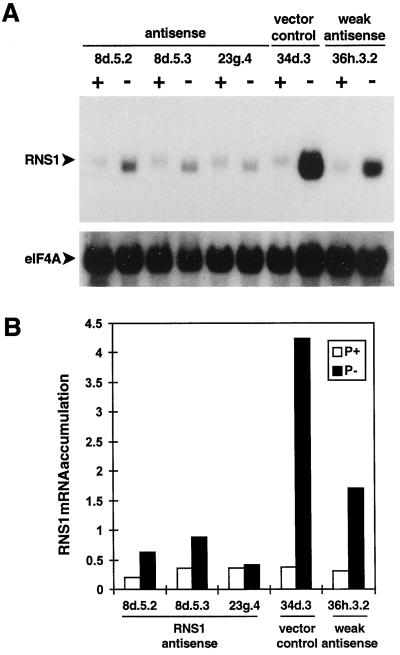
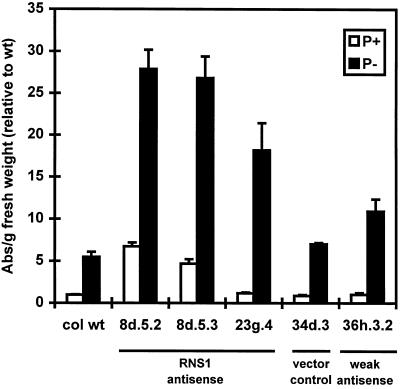
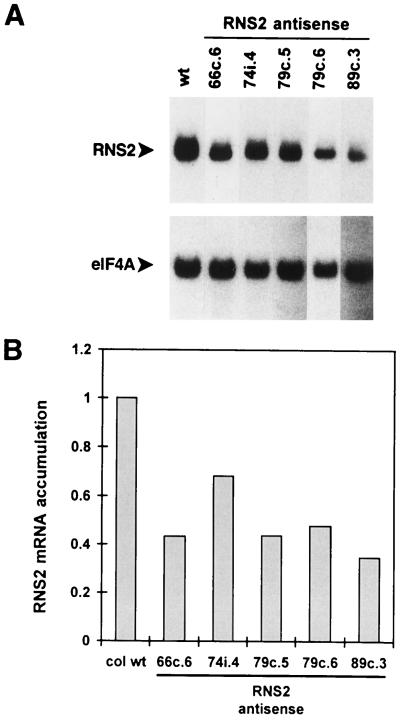
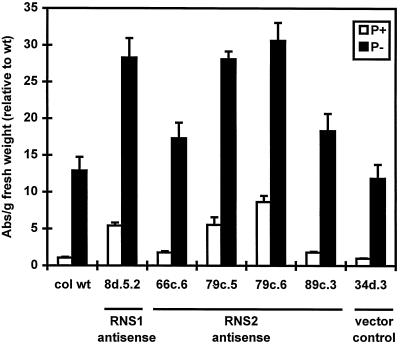
The S-like ribonucleases (RNases) RNS1 and RNS2 of Arabidopsis are members of the widespread T2 ribonuclease family, whose members also include the S-RNases, involved in gametophytic self-incompatibility in plants. Both RNS1 and RNS2 mRNAs have been shown previously to be induced by inorganic phosphate (Pi) starvation. In our study we examined this regulation at the protein level and determined the effects of diminishing RNS1 and RNS2 expression using antisense techniques. The Pi-starvation control of RNS1 and RNS2 was confirmed using antibodies specific for each protein. These specific antibodies also demonstrated that RNS1 is secreted, whereas RNS2 is intracellular. By introducing antisense constructs, mRNA accumulation was inhibited by up to 90% for RNS1 and up to 65% for RNS2. These plants contained abnormally high levels of anthocyanins, the production of which is often associated with several forms of stress, including Pi starvation. This effect demonstrates that diminishing the amounts of either RNS1 or RNS2 leads to effects that cannot be compensated for by the actions of other RNases, even though Arabidopsis contains a large number of different RNase activities. These results, together with the differential localization of the proteins, imply that RNS1 and RNS2 have distinct functions in the plant.
Figures

References
-
- Axelos M, Curie C, Mazzolini L, Bardet C, Lescure B. A protocol for transient gene expression in Arabidopsis thaliana protoplast isolated from cell suspension cultures. Plant Physiol Biochem. 1992;30:123–128.
-
- Bariola PA, Green PJ (1997) Plant ribonucleases. In G D'Alessio, JF Riordan, eds, Ribonucleases: Structures and Functions. Academic Press, New York, pp 163–190
-
- Bariola PA, Howard CJ, Taylor CB, Verburg MT, Jaglan VD, Green PJ. The Arabidopsis ribonuclease gene RNS1 is tightly controlled in response to phosphate limitation. Plant J. 1994;6:673–685. - PubMed
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci. 1993;316:1194–1199.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
