

Kainate receptor-mediated responses in the CA1 field of wild-type and GluR6-deficient mice
- PMID: 9880586
- PMCID: PMC6782223
- DOI: 10.1523/JNEUROSCI.19-02-00653.1999
Kainate receptor-mediated responses in the CA1 field of wild-type and GluR6-deficient mice
Abstract
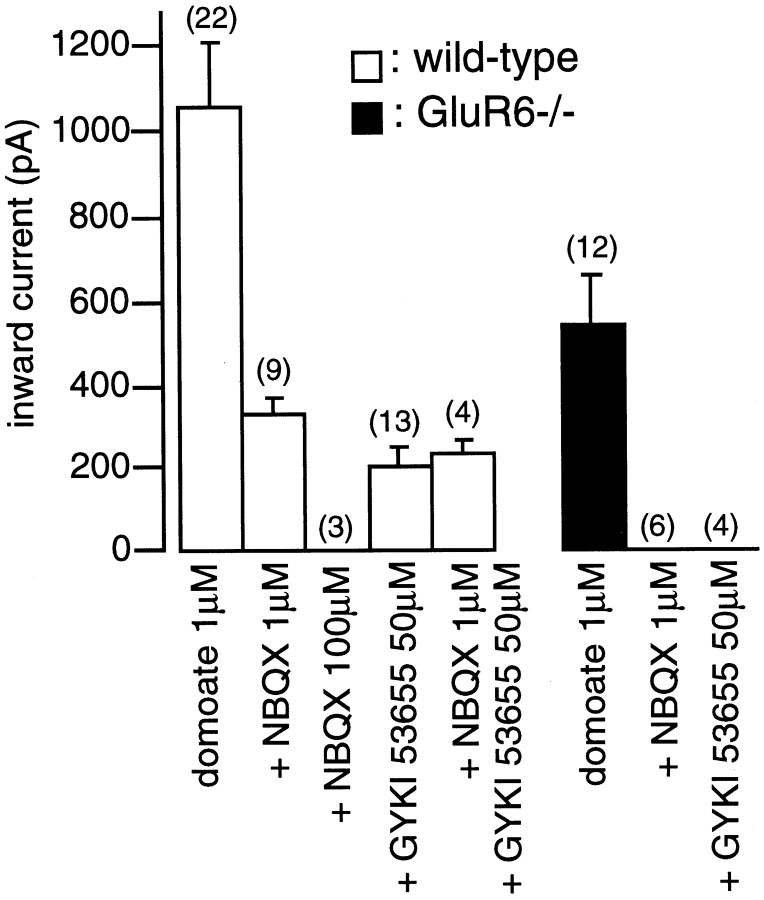
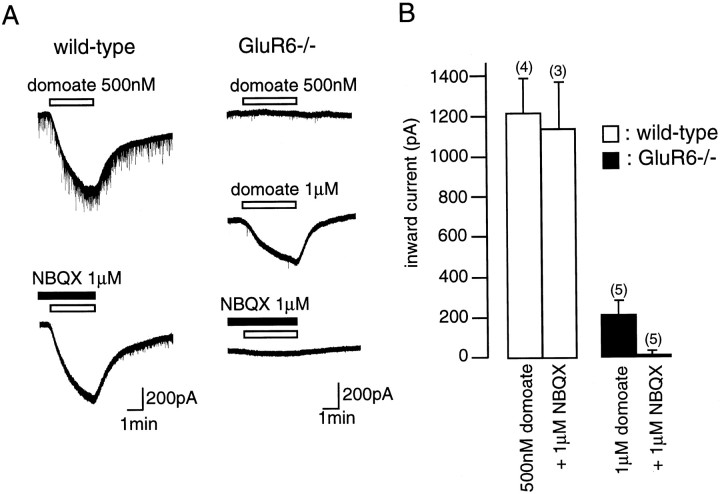
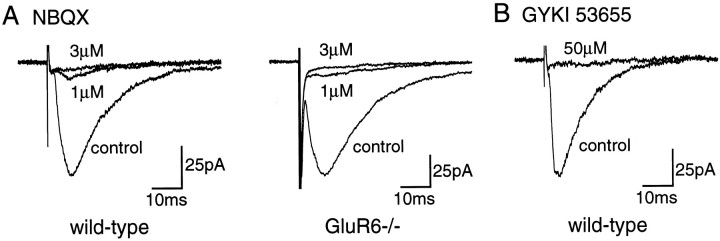
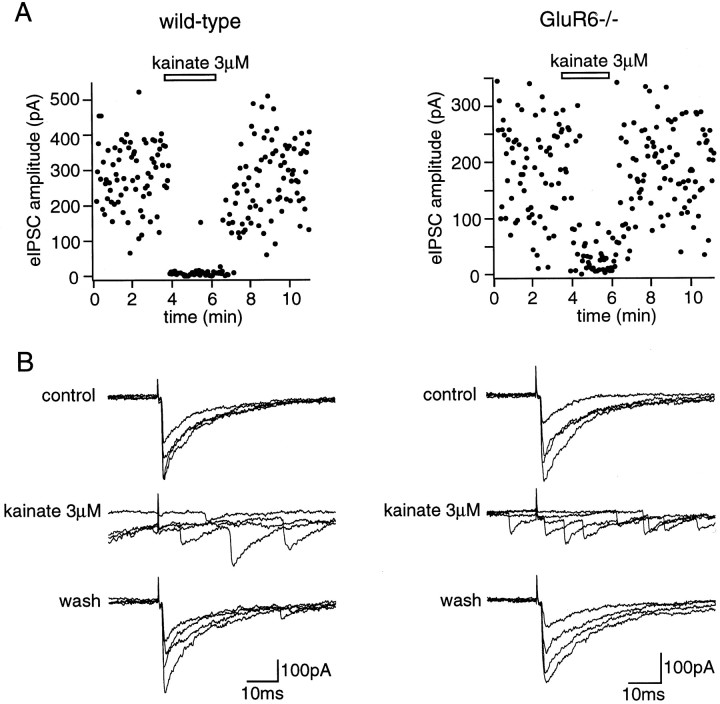
Kainate receptors are abundantly expressed in the hippocampus. Mice with disruption of kainate receptor subunits allow the genetic dissection of the role of each kainate receptor subunits in the synaptic physiology of the hippocampus, as well as in excitotoxic processes. We have compared the action of domoate and kainate on CA1 pyramidal neurons in slices from wild-type and GluR6-/- mice. The difference in the amplitude of inward currents evoked by domoate and kainate between wild-type and GluR6-/- mice demonstrates the presence of functional kainate receptors in CA1 pyramidal neurons. Block of domoate-activated inward currents by the AMPA receptor antagonists 2,3-dihydroxy-6-nitro-7-sulfonyl-benzo(F)quinoxaline (1 microM) and 1-(4-aminophenyl)-3-methylcarbamyl-4-methyl7, 8-methylenedioxy-3,4-dihydro-5H-2,3-benzodiazepine) (GYKI 53655) (50 microM) is complete in GluR6-/- mice but only partial in wild-type mice. In the presence of GYKI 53655, kainate receptor activation dramatically increases the frequency of spontaneous IPSCs in CA1 pyramidal cells from wild-type, as well as GluR6-/-, mice. This results from the kainate receptor-mediated activation of a sustained inward current and an increased action potential firing in afferent GABAergic interneurons of the CA1 field. These effects are observed in wild-type, as well as GluR6-/-, mice. Kainate receptors also decrease the amplitude of evoked IPSCs in CA1 pyramidal cells by increasing synaptic failures in wild-type and GluR6-/- mice. These results indicate that in CA1 pyramidal cells, distinct subtypes of kainate receptors mediate several functionally antagonistic effects.
Figures






References
-
- Ankri N, Legendre P, Faber DS, Korn H. Automatic detection of spontaneous synaptic responses in central neurones. J Neurosci Methods. 1994;52:87–100. - PubMed
-
- Bettler B, Mulle C. AMPA and kainate receptors. Neuropharmacology. 1995;34:123–139. - PubMed
-
- Bischoff S, Barhanin J, Bettler B, Mulle C, Heinemann S. Spatial distribution of kainate receptor subunit mRNA in the mouse basal ganglia and ventral mesencephalon. J Comp Neurol. 1997;379:541–562. - PubMed
-
- Bleakman D, Ballyk BA, Schoepp DD, Palmer AJ, Bath CP, Sharpe EF, Woolley ML, Bufton HR, Kamboj RK, Tarnawa I, Lodge D. Activity of 2,3-benzodiazepines at native rat and recombinant human glutamate receptors in vitro: stereospecificity and selectivity profiles. Neuropharmacology. 1996;35:1689–1702. - PubMed
-
- Castillo PE, Malenka RC, Nicoll RA. Kainate receptors mediate a slow postsynaptic current in hippocampal CA3 neurons. Nature. 1997;388:182–186. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous