

Characterization and cell cycle regulation of the major Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8) latent genes and their promoter
- PMID: 9882349
- PMCID: PMC103968
- DOI: 10.1128/JVI.73.2.1438-1446.1999
Characterization and cell cycle regulation of the major Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8) latent genes and their promoter
Abstract
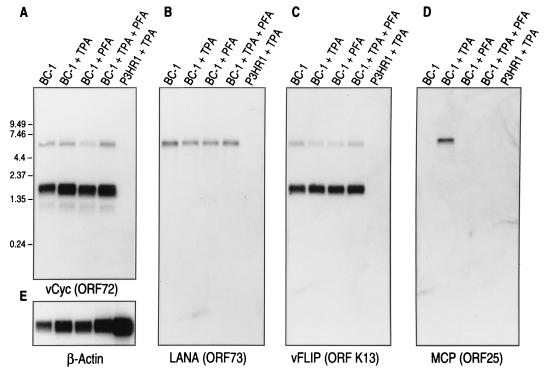
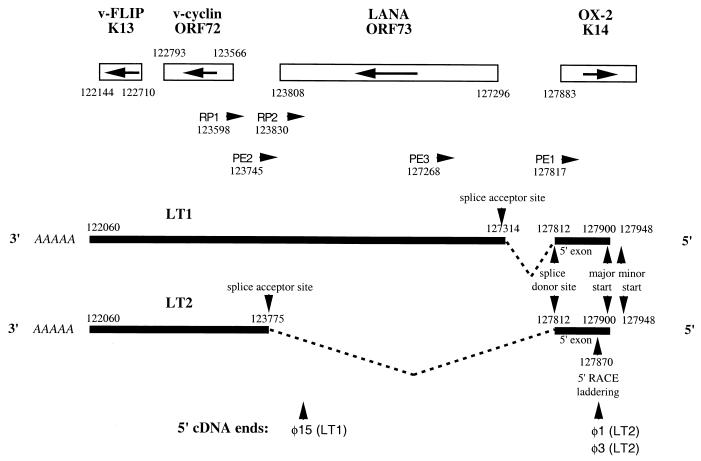
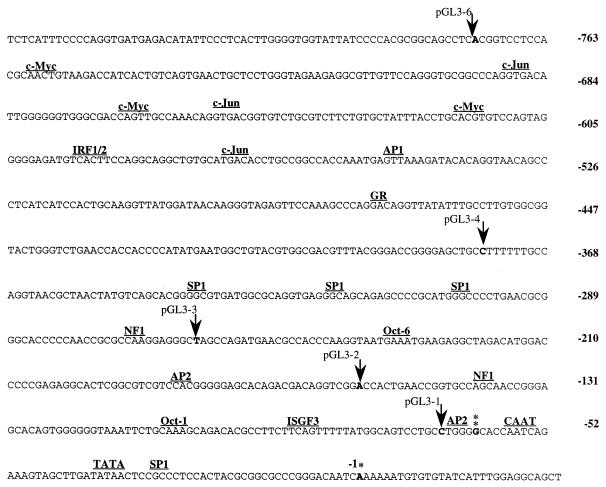
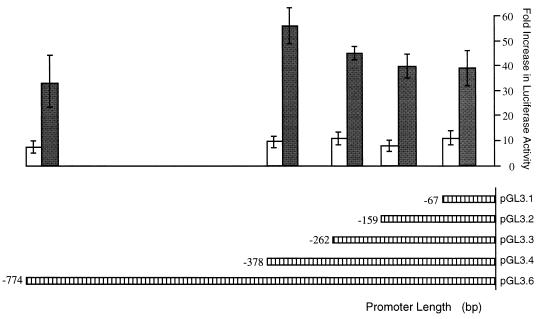
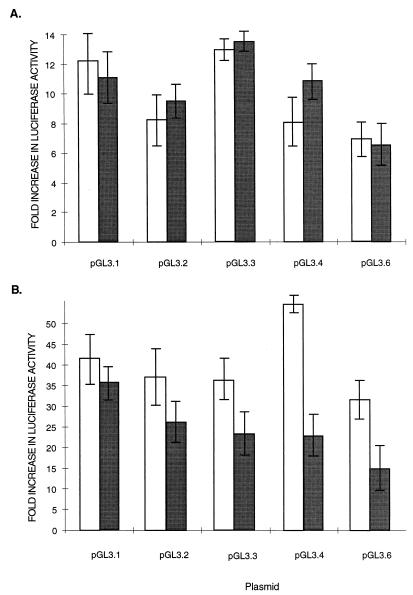
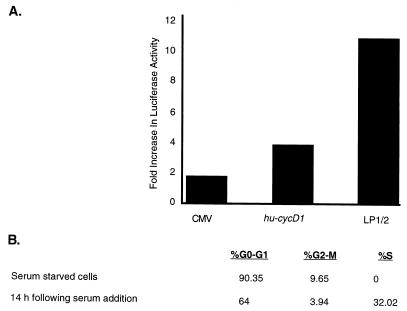
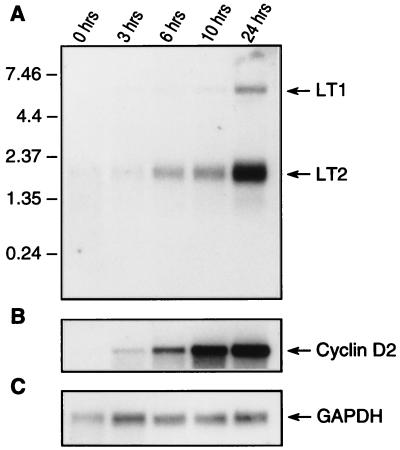
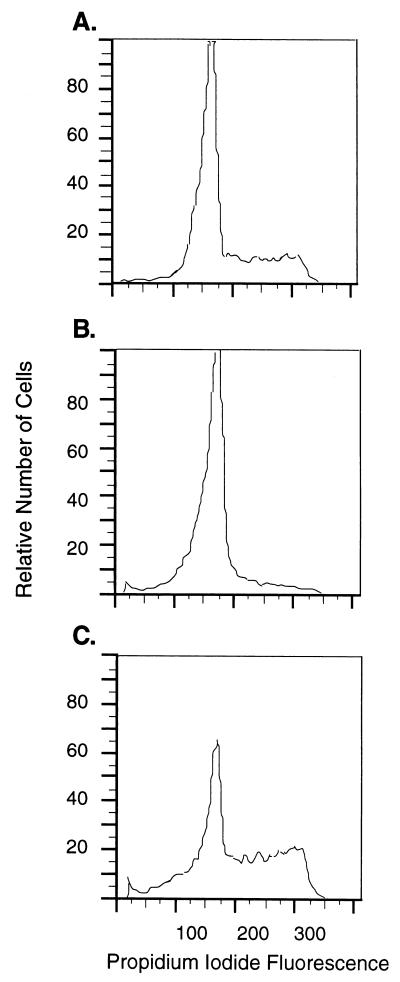
Retinoblastoma tumor suppressor protein (pRB) inhibition by tumor virus oncoproteins has been attributed to the need for these viruses to promote lytic viral nucleic acid synthesis by unscheduled entry into the S phase of the cell cycle. Kaposi's sarcoma-associated herpesvirus (KSHV or HHV8) encodes a functional cyclin (vCYC) which is expressed during latency and can direct phosphorylation of pRB. We mapped the two major latent transcripts encoding vCYC, latent transcript 1 (LT1) and LT2, by cDNA sequencing, 5' rapid amplification of cDNA ends, and primer extension analyses. Both LT1 and LT2 transcripts are spliced, originate from the same start site, and encode ORF K13 (vFLIP) as well as ORF72 (vCYC). The latency-associated nuclear antigen (LANA, ORF73) is encoded by LT1 but spliced from LT2. While differential expression of the two transcripts was not found, the promoter controlling LT1/LT2 transcription is regulated in a cell cycle-dependent manner. Activities of both KSHV LT1/LT2 and huCYC D1 luciferase promoter reporters transfected into NIH 3T3 cells increase 11- and 4-fold, respectively, after release from cell cycle arrest by serum starvation. Further, vCYC and huCYC D2 mRNA levels are low in naturally infected BCBL-1 cells arrested in late G1 with L-mimosine but increase in parallel during a 24-h period after release from cell cycle arrest. Cell cycle regulation of KSHV vCYC expression mimics cellular D cyclin regulation and may maintain infected cell cycling. This is consistent with an alternative hypothesis that tumor viruses have developed specific responses to innate cellular defenses against latent virus infection that include pRB-induced cell cycle arrest.
Figures

References
-
- Albanese C, Johnson J, Watanabe G, Eklund N, Vu D, Arnold A, Pestell R G. Transforming p21ras mutants and c-Ets-2 activate the cyclin D1 promoter through distinguishable regions. J Biol Chem. 1995;270:23589–23597. - PubMed
-
- Bates S, Phillips A C, Clark P A, Stott F, Peters G, Ludwig R L, Vousden K H. p14ARF links the tumour suppressors RB and p53. Nature. 1998;395:124–125. - PubMed
-
- Bertin J, Armstrong R C, Ottilie S, Martin D A, Wang Y, Banks S, Wang G H, Senkevich T G, Alnemri E S, Moss B, Lenardo M J, Tomaselli K J, Cohen J I. Death effector domain-containing herpesvirus and poxvirus proteins inhibit both Fas- and TNFR1-induced apoptosis. Proc Natl Acad Sci USA. 1997;94:1172–1176. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
