

Protection against a lethal avian influenza A virus in a mammalian system
- PMID: 9882351
- PMCID: PMC103970
- DOI: 10.1128/JVI.73.2.1453-1459.1999
Protection against a lethal avian influenza A virus in a mammalian system
Abstract
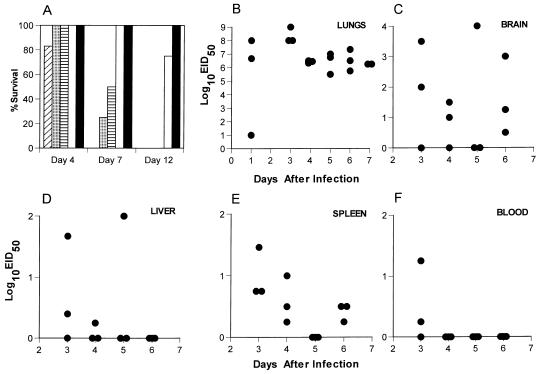
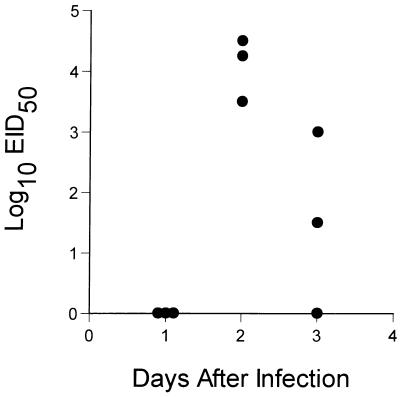
The question of how best to protect the human population against a potential influenza pandemic has been raised by the recent outbreak caused by an avian H5N1 virus in Hong Kong. The likely strategy would be to vaccinate with a less virulent, laboratory-adapted H5N1 strain isolated previously from birds. Little attention has been given, however, to dissecting the consequences of sequential exposure to serologically related influenza A viruses using contemporary immunology techniques. Such experiments with the H5N1 viruses are limited by the potential risk to humans. An extremely virulent H3N8 avian influenza A virus has been used to infect both immunoglobulin-expressing (Ig+/+) and Ig-/- mice primed previously with a laboratory-adapted H3N2 virus. The cross-reactive antibody response was very protective, while the recall of CD8(+) T-cell memory in the Ig-/- mice provided some small measure of resistance to a low-dose H3N8 challenge. The H3N8 virus also replicated in the respiratory tracts of the H3N2-primed Ig+/+ mice, generating secondary CD8(+) and CD4(+) T-cell responses that may contribute to recovery. The results indicate that the various components of immune memory operate together to provide optimal protection, and they support the idea that related viruses of nonhuman origin can be used as vaccines.
Figures
References
-
- Allan W, Tabi Z, Clearly A, Doherty P C. Cellular events in the lymph node and lung of mice with influenza: consequences of depleting CD4+ T cells. J Immunol. 1990;144:3980–3986. - PubMed
-
- Altman J D, Moss P A H, Goulder P J R, Barouch D H, McHeyzer-Williams M G, Bell J I, McMichael A J, Davis M M. Phenotypic analysis of antigen-specific T lymphocytes. Science. 1996;274:94–96. - PubMed
-
- Claas E C J, Osterhaus A D M E, van Beek R, De Jong J C, Rimmelzwaan G F, Senne D A, Karauss S, Shortridge K F, Webster R G. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet. 1998;351:472–477. - PubMed
-
- Cohen J. The flu pandemic that might have been. Science. 1997;277:1600–1601. - PubMed
-
- Deckhut A M, Allan W, McMickle A, Eichelberger M, Blackman M A, Doherty P C, Woodland D L. Prominent use of Vβ8.3 T cells in H-2Db nucleoprotein epitope. J Immunol. 1993;151:2658–2666. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
