

doi: 10.1128/JVI.73.2.1624-1629.1999.
Visualization of protein-RNA interactions in cytoplasmic polyhedrosis virus
Affiliations
- PMID: 9882369
- PMCID: PMC103988
- DOI: 10.1128/JVI.73.2.1624-1629.1999
Item in Clipboard
Visualization of protein-RNA interactions in cytoplasmic polyhedrosis virus
J Virol.
1999 Feb.
Abstract
Unlike the multiple-shelled organization of other Reoviridae members, cytoplasmic polyhedrosis virus (CPV) has a single-shelled capsid. The three-dimensional structures of full and empty CPV by electron cryomicroscopy show identical outer shells but differ inside. The outer surface reveals a T=1 icosahedral shell decorated with spikes at its icosahedral vertices. The internal space of the empty CPV is unoccupied except for 12 mushroom-shaped densities attributed to the transcriptional enzyme complexes. The ordered double-stranded RNA inside the full capsid forms spherical shells spaced 25 A apart. The RNA-protein interactions suggest a mechanism for RNA transcription and release.
Figures
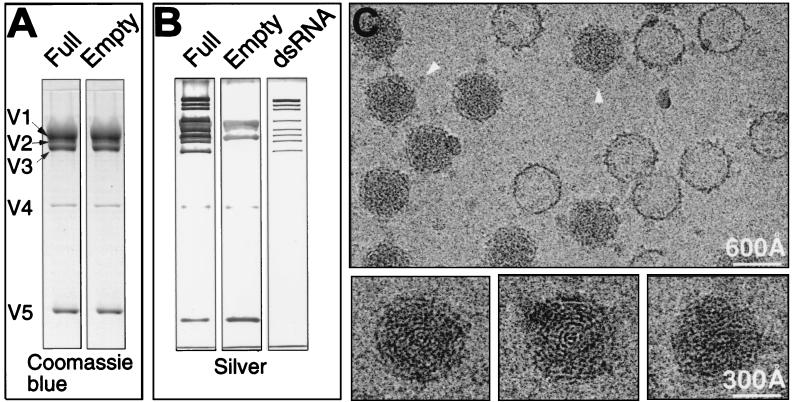
SDS-PAGE analyses and electron cryomicroscopic images of purified CPV capsids. Fifth-instar larvae of B. mori were infected with CPV by spraying a suspension of polyhedra (108/ml) onto mulberry leaves. Ten days after infection, midguts were removed from the larvae and polyhedra were purified by a method modified from that of Hayashi and Bird (11). In our procedure, 0.5% SDS and 100 μg of trypsin/ml were used instead of the 1% sodium deoxycholate and an enzyme mixture (RNase A, DNase, trypsin [50 μg/ml each]). We also used 20 mM phosphate-buffered saline (PBS) buffer (pH 7.4) instead of TK buffer (0.03 M tris-HCl buffer, pH 7.5; 0.025M KCl). The polyhedra were resuspended in 0.2 M sodium carbonate-sodium bicarbonate buffer. After 1 h at 30°C, the pH was adjusted to 7.4 with 20 mM NaH2PO4 and the mixture was centrifuged at 10,000 × g for 10 min. The virus particles were pelleted from the supernatant at 90,000 × g for 70 min. The pellet was resuspended in PBS and purified in 4 ml of 10 to 40% (wt/wt) sucrose gradient by centrifugation at 55,000 × g for 1 h. Virus-containing fractions were recovered. After being diluted with PBS, the viral sample was pelleted again at 90,000 × g for 70 min and resuspended in PBS. (A) SDS-PAGE analyses of the purified full and empty capsids were carried out with Coomassie blue staining to identify the differences of protein compositions between the full and empty capsids. Viral proteins V1 to V5 are labeled. (B) The SDS gels stained with silver reveal the nucleic acid compositions as well as protein compositions in full and empty capsids. The right panel shows the profile of the RNA genome extracted from the full capsid. (C) (Upper panel) Typical area of a close-to-focus electron micrograph (1.46 μm defocus) in a focal pair of ice-embedded CPV capsids taken at 100 kV. The micrographs were digitized on a Zeiss SCAI microdensitometer (Carl Zeiss, Inc., Englewood, Colo.) at a step size of 4.67 Å/pixel, and individual virus particles were extracted into individual images of 200 × 200 pixels. Arrowheads indicate the A spikes visible in some capsid views. (Lower panels) three different full capsids are enlarged to show the characteristic fingerprint patterns.
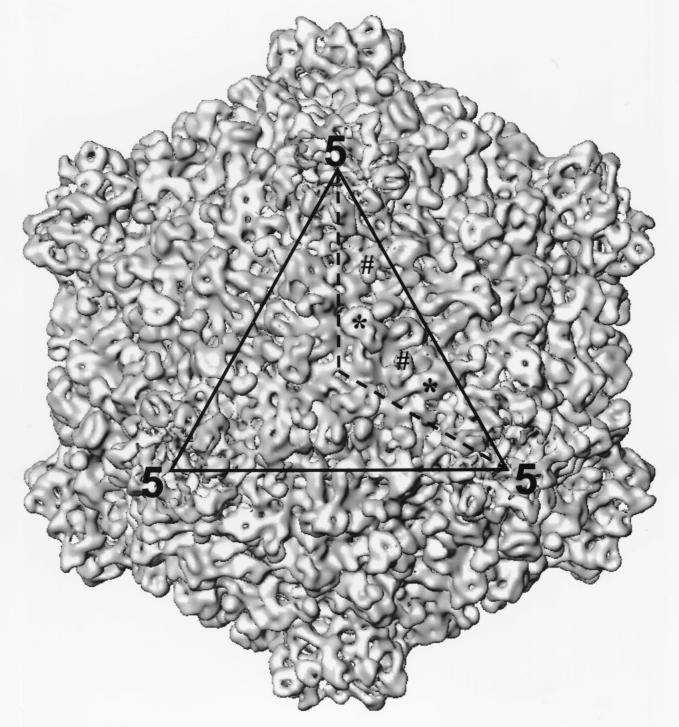
Surface representation of the 3D structure of the full CPV capsid at 17-Å resolution as viewed along the icosahedral threefold axis. One of the three asymmetric units outlined in the icosahedral triangular face (dotted line) contains two SPs and two LPs which are designated by # and ∗, respectively.
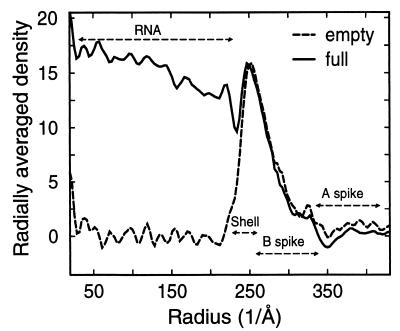
The averaged density distribution of the final 3D reconstructions of the full and empty capsids as a function of particle radius. The density beyond a radius of 230 Å appears almost identical in the empty and full capsids. Below a radius of 230 Å, there are several layers of high density in the full capsid, which are completely absent in the empty capsid. The radial range locations of structural components in full and empty capsids are indicated.
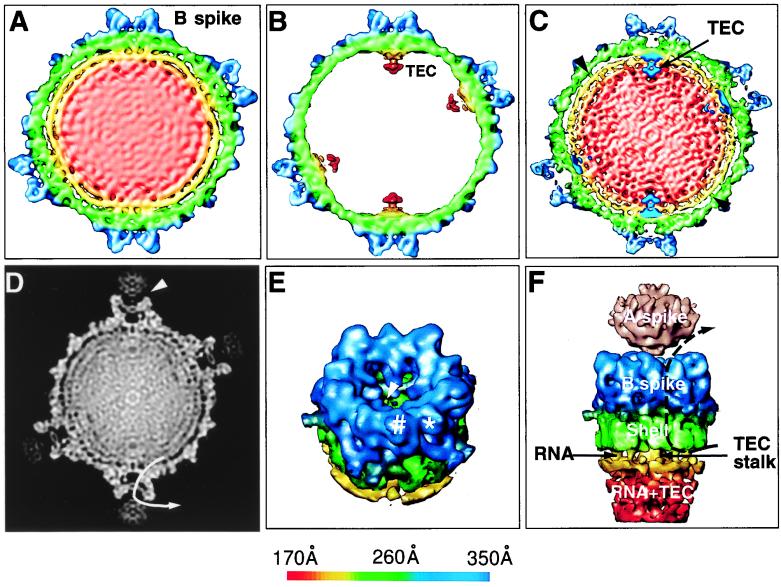
Visualization of protein-RNA interactions in CPV. (A and B) Structural comparison of full and empty capsids. A central 50-Å thick slice was extracted from the 3D reconstructions of the full (A) and empty (B) capsids at 25-Å resolution. Both maps were contoured at 1ς above the mean except for the internal complexes of the empty capsid, where 0.5ς was used to reveal the TEC. (C) A 50-Å thick central slice from the 17-Å full capsid reconstruction was superimposed with the TECs extracted from the reconstruction of the empty capsids. The interactions between the capsid proteins and the RNA were strongest at the base of the spike. Minor interactions were also observed close to the twofold axis (arrowhead). The densities attributed to dsRNA inside the full capsid formed two shells between the 160- to 230-Å radii, with a distance of 25 Å between adjacent RNA shells. (D) A 3-Å-thick slice extracted from the center of the 3D reconstruction of the full capsid is displayed in gray scale. The A spike is loosely connected to the B spike. The A spike has a globular structure, whereas the B spike is a hollow protrusion from the shell. The pathway of one of the five channels, which connects the inside and outside the capsid, is depicted by the arrow line. (E) The B spike and a portion of the viral shell were extracted to reveal the hole at the top cover of the B spike cavity and the terminal end of the channel connecting the inside and outside of the capsid (arrowhead). Each of the five subunits of the spike has two discernible subdomains (∗ and #). (F) A portion of the full map at the icosahedral vertex was extracted and viewed from the side to illustrate the interactions among the RNA, the B spike, and the shell. The A spike, visible only when displayed at 0.5ς above the mean, is shown in pink. The channel connecting the inside with the outside indicated in panel D is illustrated by the dotted line. Except for the A spike in panel F, the maps are color coded according to the radius of the capsid, using the scheme shown at the bottom.
References
-
- Crowther R A. Procedures for three-dimensional reconstruction of spherical viruses by Fourier synthesis from electron micrographs. Phil Trans R Soc Lond B. 1971;261:221–230. - PubMed
-
- Dai R M, Wu A Z, Sun Y K. The protein subunits of the double-stranded RNA dependent RNA polymerase and methyltransferase of the cytoplasmic polyhedrosis virus of silkworm, Bombyx mori. Sci Sin Ser B (Chem Biol Agric Med Earth Sci) 1986;29:1267–1272. - PubMed
-
- Dryden K A, Farsetta D L, Wang G, Keegan J M, Fields B N, Baker T S, Nibert M L. Internal structures containing transcriptase-related proteins in top component particles of mammalian orthoreovirus. Virology. 1998;245:33–46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
