

Heat-induced synthesis of sigma32 in Escherichia coli: structural and functional dissection of rpoH mRNA secondary structure
- PMID: 9882652
- PMCID: PMC93392
- DOI: 10.1128/JB.181.2.401-410.1999
Heat-induced synthesis of sigma32 in Escherichia coli: structural and functional dissection of rpoH mRNA secondary structure
Abstract
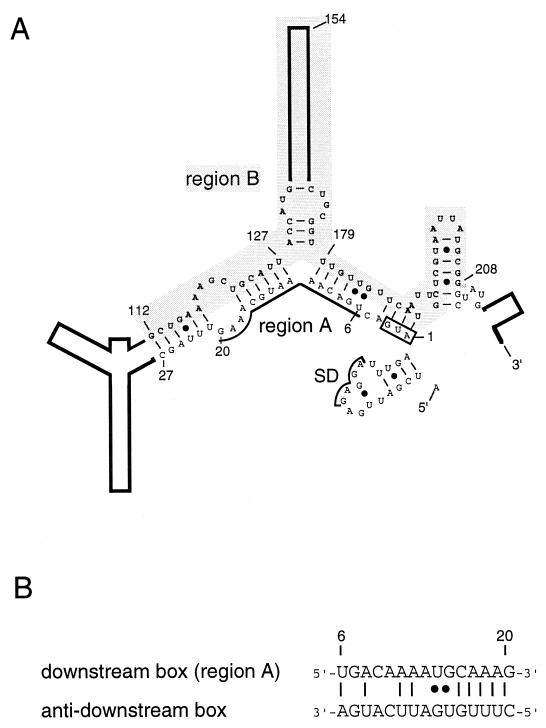
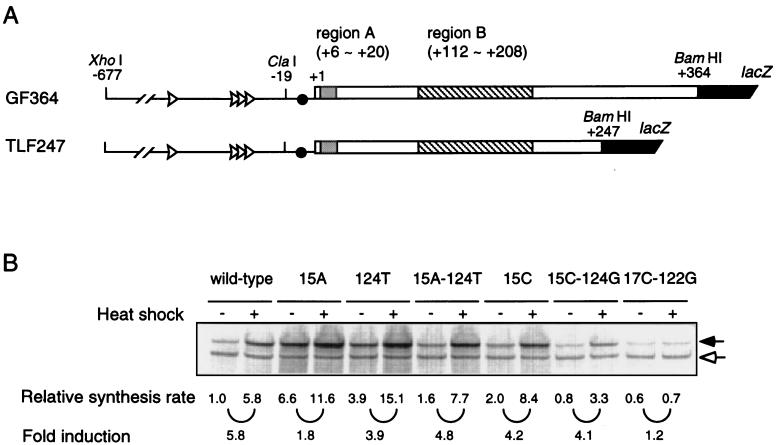
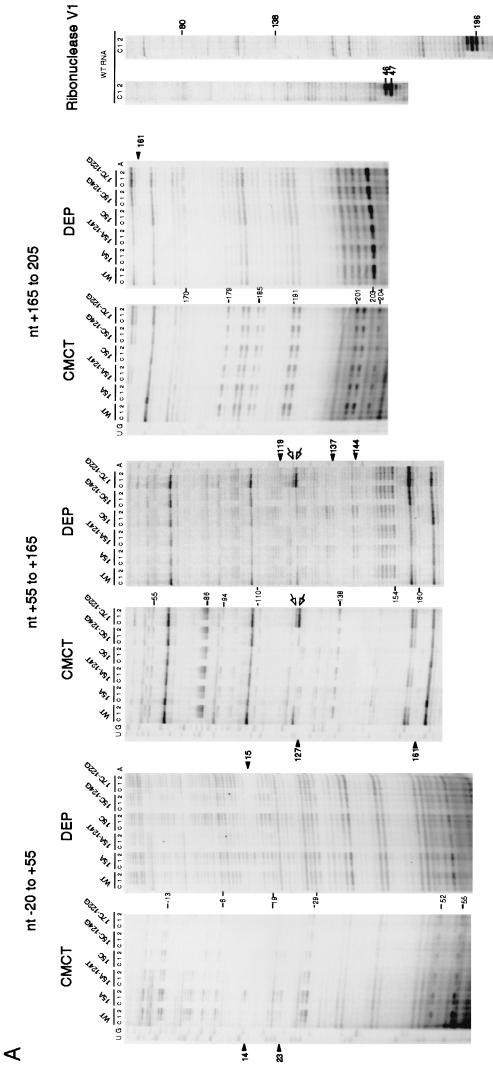
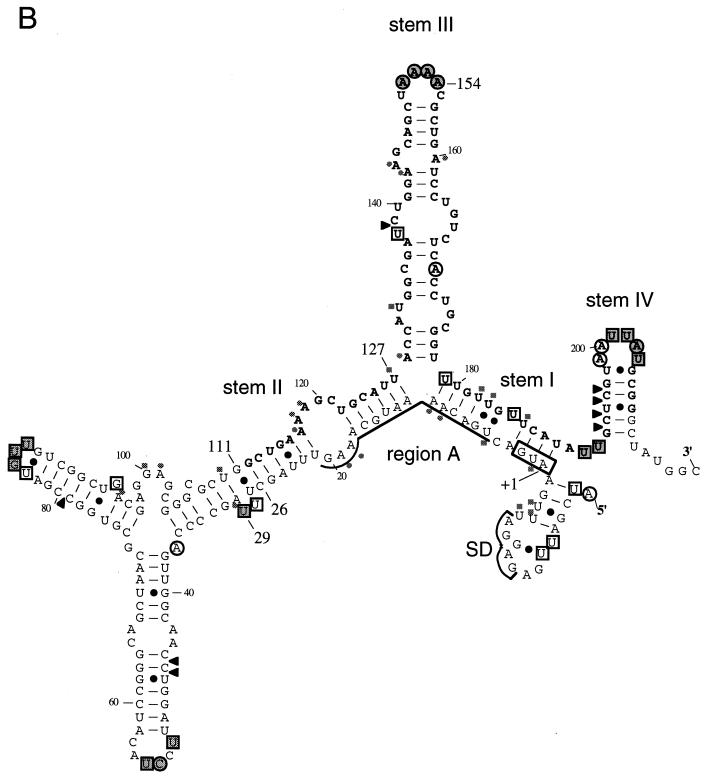
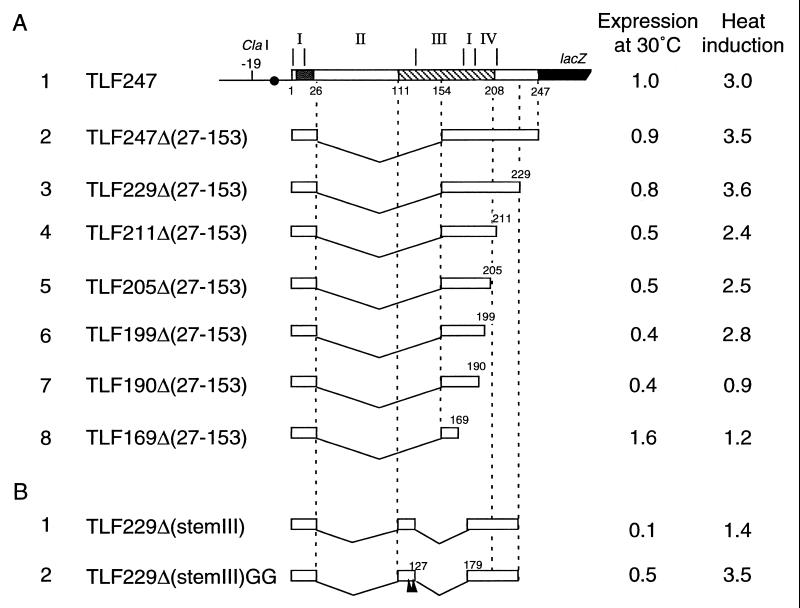
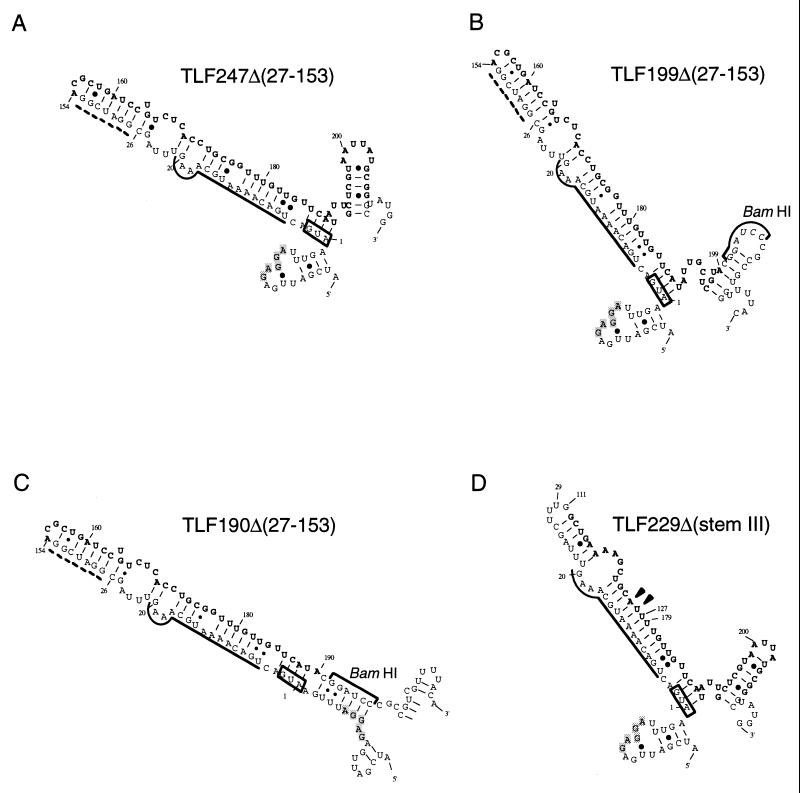
The heat shock response in Escherichia coli depends primarily on the increased synthesis and stabilization of otherwise scarce and unstable sigma32 (rpoH gene product), which is required for the transcription of heat shock genes. The heat-induced synthesis of sigma32 occurs at the level of translation, and genetic evidence has suggested the involvement of a secondary structure at the 5' portion (nucleotides -19 to +247) of rpoH mRNA in regulation. We now present evidence for the mRNA secondary structure model by means of structure probing of RNA with chemical and enzymatic probes. A similar analysis of several mutant RNAs with a mutation predicted to alter a base pairing or with two compensatory mutations revealed altered secondary structures consistent with the expression and heat inducibility of the corresponding fusion constructs observed in vivo. These findings led us to assess the possible roles of each of the stem-loop structures by analyzing an additional set of deletions and base substitutions. The results indicated not only the primary importance of base pairings between the translation initiation region of ca. 20 nucleotides (the AUG initiation codon plus the "downstream box") and the internal region of rpoH mRNA but also the requirement of appropriate stability of mRNA secondary structures for characteristic thermoregulation, i.e., repression at a low temperature and induction upon a temperature upshift.
Figures
References
-
- Bukau B. Regulation of the Escherichia coli heat shock response. Mol Microbiol. 1993;9:671–680. - PubMed
-
- Casadaban M J. Transposition and fusion of the lac genes to selected promoters in Escherichia coli using bacteriophage lambda and Mu. J Mol Biol. 1976;104:541–555. - PubMed
-
- Christiansen J, Egebjerg J, Larsen N, Garrett R. Analysis of rRNA structure: experimental and theoretical considerations. In: Spedding G, editor. Ribosome and protein synthesis: a practical approach. Oxford, England: IRL Press; 1991. pp. 229–252.
-
- Craig E A, Gross C A. Is hsp70 the cellular thermometer? Trends Biochem Sci. 1991;16:135–140. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
