

Subunit II of Bacillus subtilis cytochrome c oxidase is a lipoprotein
- PMID: 9882689
- PMCID: PMC93429
- DOI: 10.1128/JB.181.2.685-688.1999
Subunit II of Bacillus subtilis cytochrome c oxidase is a lipoprotein
Abstract
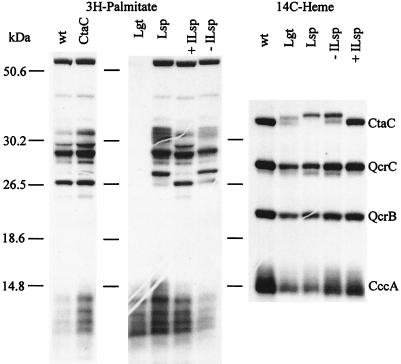
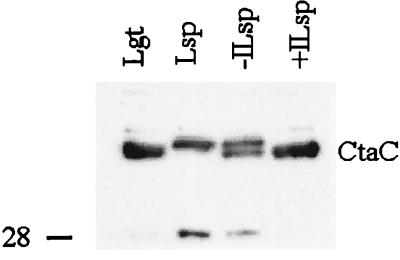
The sequence of the N-terminal end of the deduced ctaC gene product of Bacillus species has the features of a bacterial lipoprotein. CtaC is the subunit II of cytochrome caa3, which is a cytochrome c oxidase. Using Bacillus subtilis mutants blocked in lipoprotein synthesis, we show that CtaC is a lipoprotein and that synthesis of the membrane-bound protein and covalent binding of heme to the cytochrome c domain is not dependent on processing at the N-terminal part of the protein. Mutants blocked in prolipoprotein diacylglyceryl transferase (Lgt) or signal peptidase type II (Lsp) are, however, deficient in cytochrome caa3 enzyme activity. Removal of the signal peptide from the CtaC polypeptide, but not lipid modification, is seemingly required for formation of functional enzyme.
Figures

References
-
- Calhoun M W, Thomas J W, Gennis R B. The cytochrome oxidase superfamily of redox-driven proton pumps. Trends Biochem Sci. 1994;19:325–330. - PubMed
-
- Chamberlain J P. Fluorographic detection of radioactivity in polyacrylamide gels with the water-soluble fluor, sodium salicylate. Anal Biochem. 1979;98:132–135. - PubMed
-
- Dartois V, Djavakhishvili T, Hoch J A. KapB is a lipoprotein required for KinB signal transduction and activation of the phosphorelay to sporulation in Bacillus subtilis. Mol Microbiol. 1997;26:1097–1108. - PubMed
-
- Hayashi S, Wu H C. Identification and characterization of lipid-modified proteins in bacteria. In: Hooper N M, Turner A J, editors. Lipid modification of proteins. Oxford, England: IRL Press; 1992. pp. 261–285.
-
- Hederstedt L. Molecular properties, genetics, and biosynthesis of Bacillus subtilis succinate dehydrogenase complex. Methods Enzymol. 1986;126:399–414. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
