

A complex containing RNA polymerase II, Paf1p, Cdc73p, Hpr1p, and Ccr4p plays a role in protein kinase C signaling
- PMID: 9891041
- PMCID: PMC116036
- DOI: 10.1128/MCB.19.2.1056
A complex containing RNA polymerase II, Paf1p, Cdc73p, Hpr1p, and Ccr4p plays a role in protein kinase C signaling
Abstract
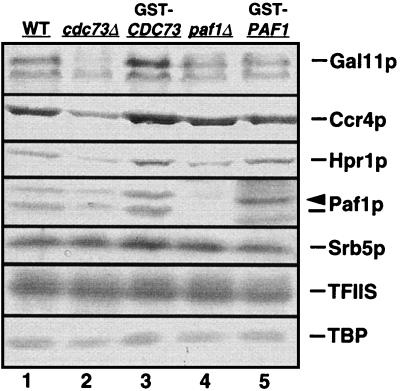
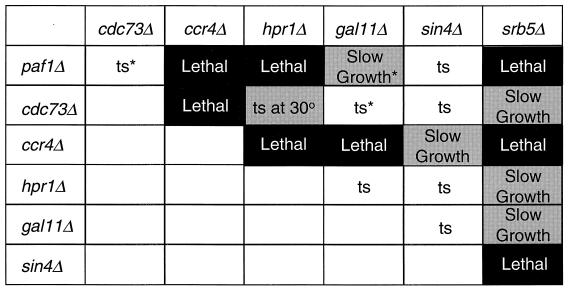
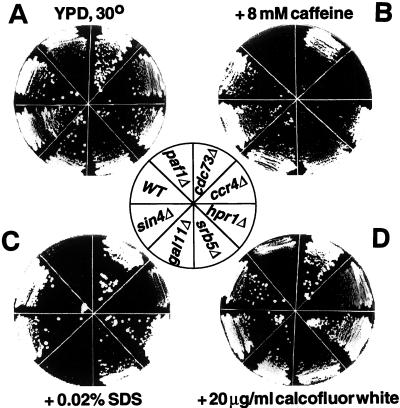
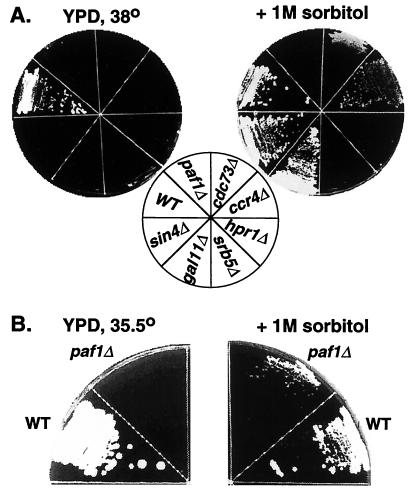
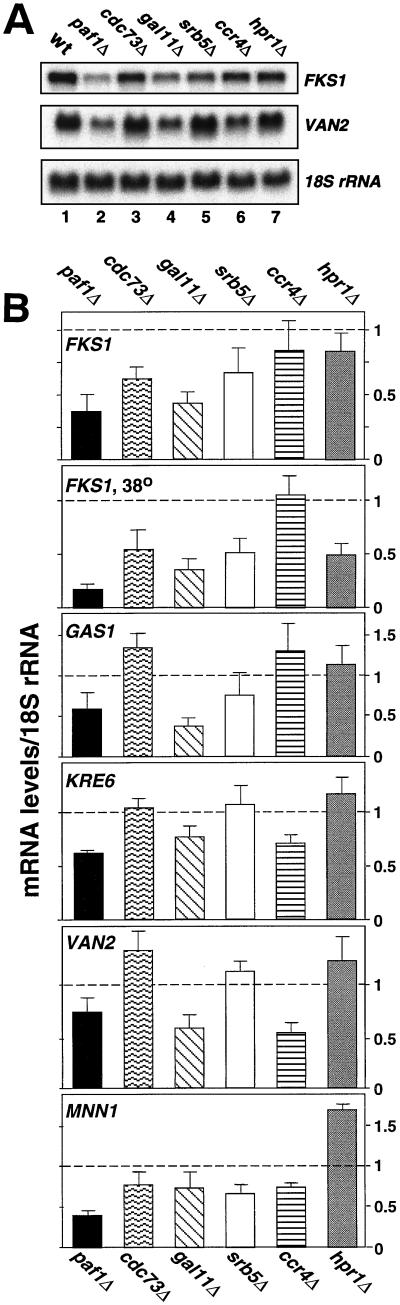
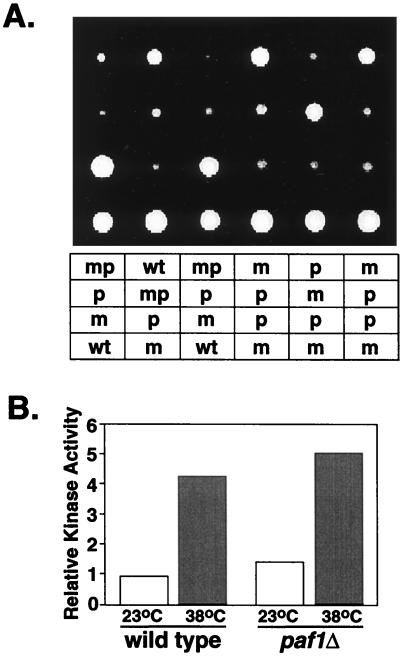
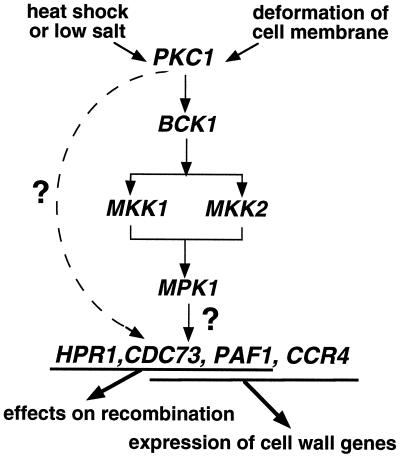
Yeast contains at least two complex forms of RNA polymerase II (Pol II), one including the Srbps and a second biochemically distinct form defined by the presence of Paf1p and Cdc73p (X. Shi et al., Mol. Cell. Biol. 17:1160-1169, 1997). In this work we demonstrate that Ccr4p and Hpr1p are components of the Paf1p-Cdc73p-Pol II complex. We have found many synthetic genetic interactions between factors within the Paf1p-Cdc73p complex, including the lethality of paf1Delta ccr4Delta, paf1Delta hpr1Delta, ccr4Delta hpr1Delta, and ccr4Delta gal11Delta double mutants. In addition, paf1Delta and ccr4Delta are lethal in combination with srb5Delta, indicating that the factors within and between the two RNA polymerase II complexes have overlapping essential functions. We have used differential display to identify several genes whose expression is affected by mutations in components of the Paf1p-Cdc73p-Pol II complex. Additionally, as previously observed for hpr1Delta, deleting PAF1 or CDC73 leads to elevated recombination between direct repeats. The paf1Delta and ccr4Delta mutations, as well as gal11Delta, demonstrate sensitivity to cell wall-damaging agents, rescue of the temperature-sensitive phenotype by sorbitol, and reduced expression of genes involved in cell wall biosynthesis. This unusual combination of effects on recombination and cell wall integrity has also been observed for mutations in genes in the Pkc1p-Mpk1p kinase cascade. Consistent with a role for this novel form of RNA polymerase II in the Pkc1p-Mpk1p signaling pathway, we find that paf1Delta mpk1Delta and paf1Delta pkc1Delta double mutants do not demonstrate an enhanced phenotype relative to the single mutants. Our observation that the Mpk1p kinase is fully active in a paf1Delta strain indicates that the Paf1p-Cdc73p complex may function downstream of the Pkc1p-Mpk1p cascade to regulate the expression of a subset of yeast genes.
Figures


References
-
- Botstein D, Falco S C, Stewart S E, Brennan M, Scherer S, Stinchcomb D T, Struhl K, Davis R W. Sterile host yeasts (SHY): a eukaryotic system of biological containment for recombinant DNA experiments. Gene. 1979;8:17–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases