

The SWI/SNF complex creates loop domains in DNA and polynucleosome arrays and can disrupt DNA-histone contacts within these domains
- PMID: 9891080
- PMCID: PMC116075
- DOI: 10.1128/MCB.19.2.1470
The SWI/SNF complex creates loop domains in DNA and polynucleosome arrays and can disrupt DNA-histone contacts within these domains
Abstract
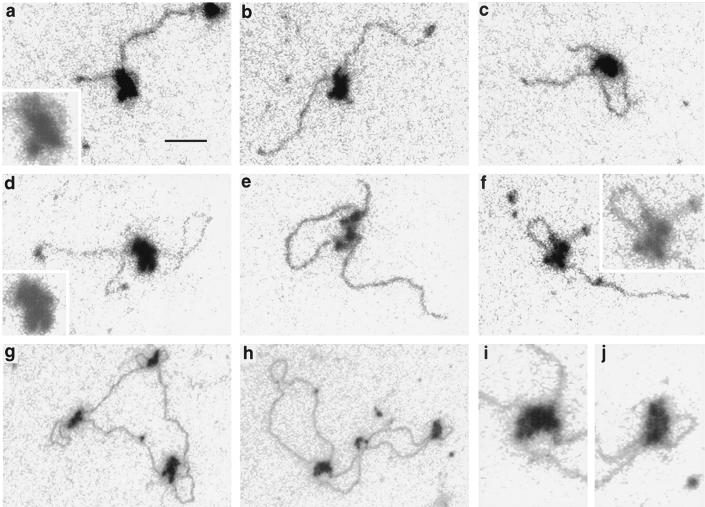
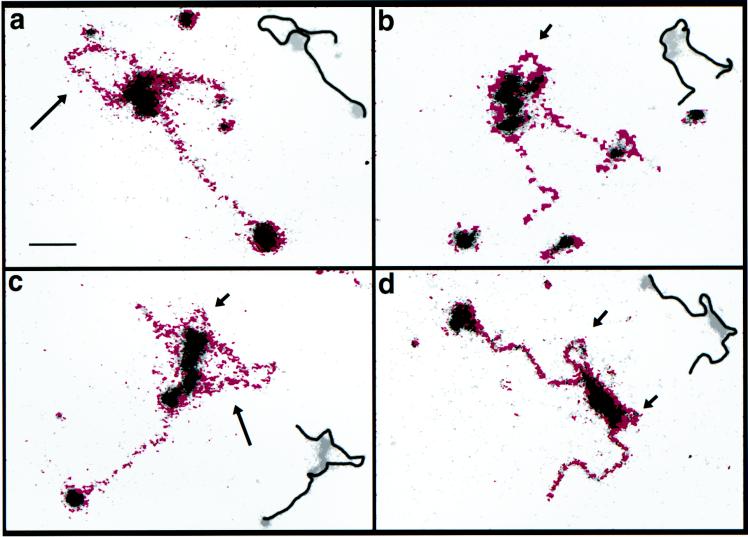
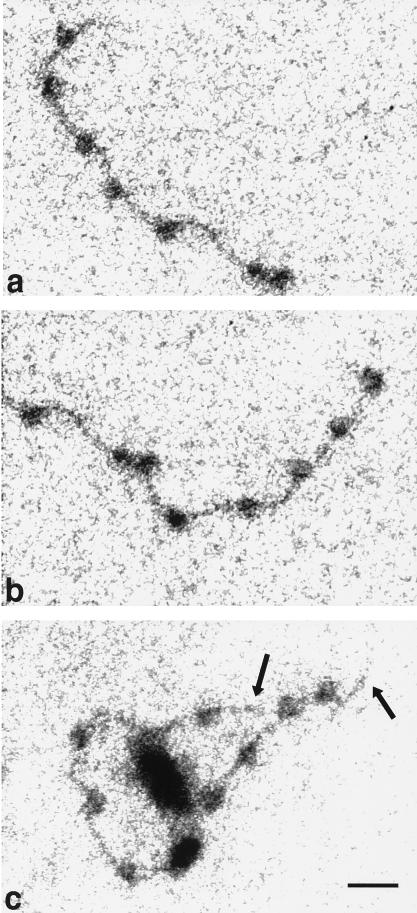
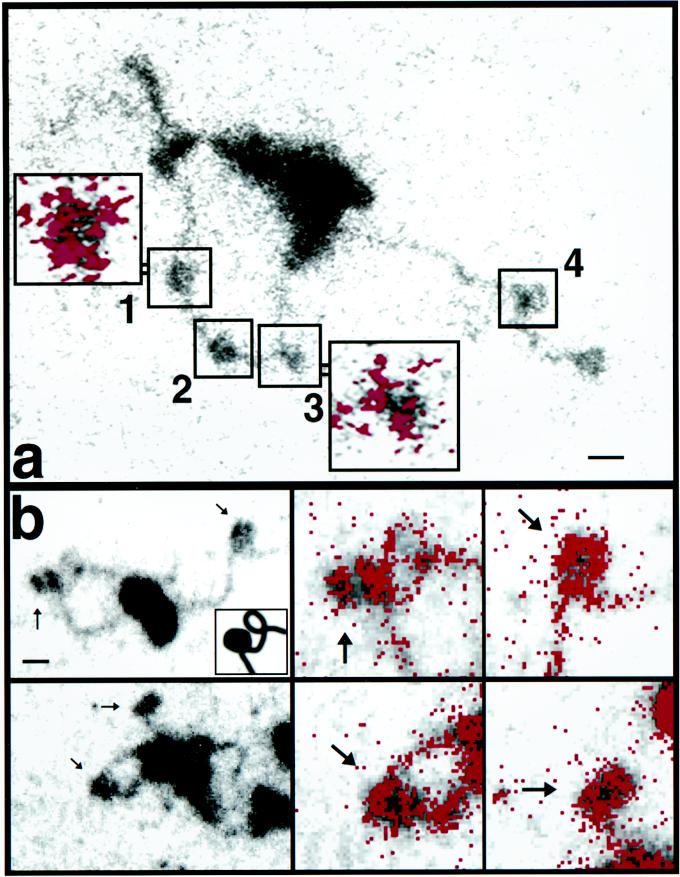
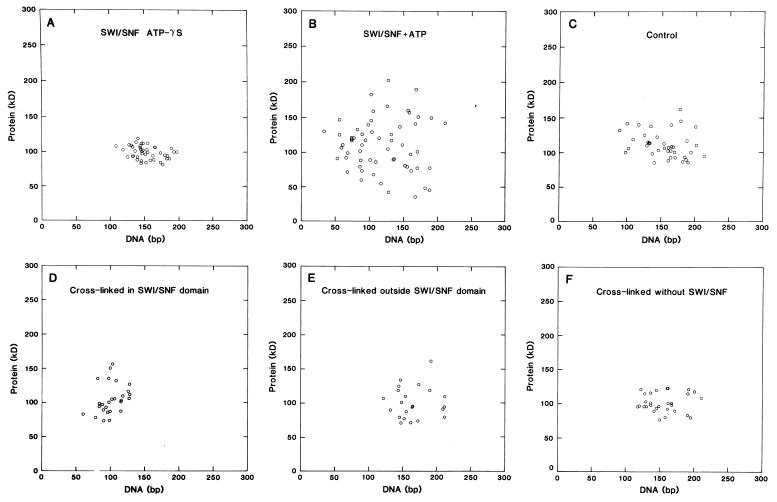
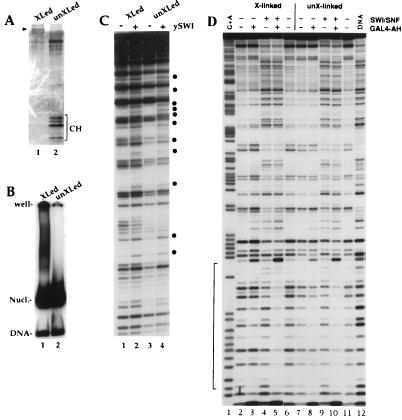
To understand the mechanisms by which the chromatin-remodeling SWI/SNF complex interacts with DNA and alters nucleosome organization, we have imaged the SWI/SNF complex with both naked DNA and nucleosomal arrays by using energy-filtered microscopy. By making ATP-independent contacts with DNA at multiple sites on its surface, SWI/SNF creates loops, bringing otherwise-distant sites into close proximity. In the presence of ATP, SWI/SNF action leads to the disruption of nucleosomes within domains that appear to be topologically constrained by the complex. The data indicate that the action of one SWI/SNF complex on an array of nucleosomes can lead to the formation of a region where multiple nucleosomes are disrupted. Importantly, nucleosome disruption by SWI/SNF results in a loss of DNA content from the nucleosomes. This indicates a mechanism by which SWI/SNF unwraps part of the nucleosomal DNA.
Figures
References
-
- Bazett-Jones D P. Empirical basis for phosphorus mapping and structure determination of DNA: protein complexes by electron spectroscopic imaging. Microbeam Anal. 1993;2:69–79.
-
- Bazett-Jones D P, Leblanc B, Herfort M, Moss T. Short-range DNA looping by the Xenopus HMG-box transcription factor, xUBF. Science. 1994;264:1134–1137. - PubMed
-
- Cairns B R, Lorch Y, Li Y, Zhang M, Lacomis L, Erdjument-Bromage H, Tempst P, Du J, Laurent B, Kornberg R D. RSC, an essential, abundant chromatin re-modeling complex. Cell. 1996;87:1249–1260. - PubMed
-
- Carlson M, Laurent B C. The SWI/SNF family of global transcriptional activators. Curr Opin Cell Biol. 1994;6:396–402. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases