

Involvement of regulatory and catalytic subunits of phosphoinositide 3-kinase in NF-kappaB activation
- PMID: 9892650
- PMCID: PMC15153
- DOI: 10.1073/pnas.96.2.429
Involvement of regulatory and catalytic subunits of phosphoinositide 3-kinase in NF-kappaB activation
Abstract
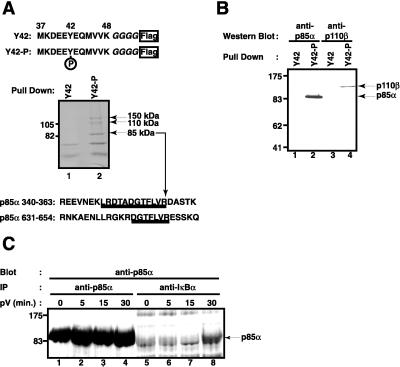
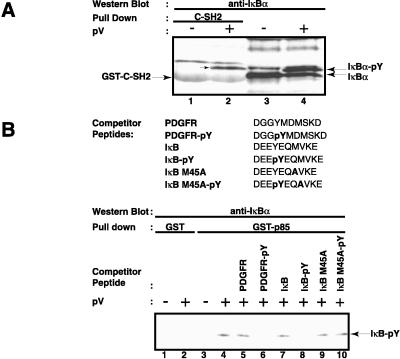
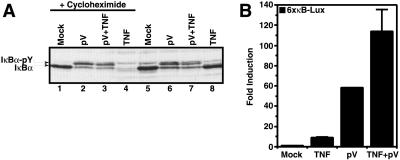
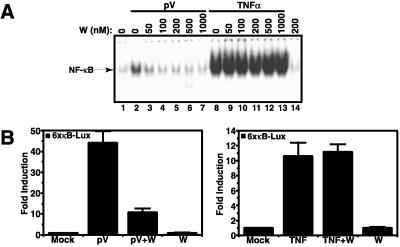
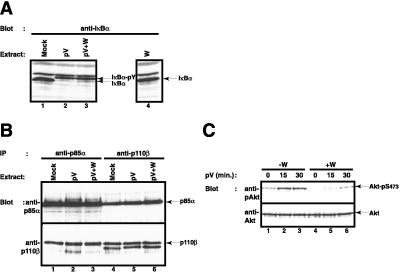
Hypoxia, reoxygenation, and the tyrosine phosphatase inhibitor pervanadate activate the transcription factor NF-kappaB, involving phosphorylation of its inhibitor IkappaB-alpha on tyrosine 42. This modification does not lead to degradation of IkappaB by the proteasome/ubiquitin pathway, as is seen on stimulation of cells with proinflammatory cytokines. It is currently unknown how tyrosine-phosphorylated IkappaB is removed from NF-kappaB. Here we show that p85alpha, the regulatory subunit of PI3-kinase, specifically associates through its Src homology 2 domains with tyrosine-phosphorylated IkappaB-alpha in vitro and in vivo after stimulation of T cells with pervanadate. This association could provide a mechanism by which newly tyrosine-phosphorylated IkappaB is sequestered from NF-kappaB. Another mechanism by which PI3-kinase contributed to NF-kappaB activation in response to pervanadate appeared to involve its catalytic p110 subunit. This was evident from the inhibition of pervanadate-induced NF-kappaB activation and reporter gene induction by treatment of cells with nanomolar amounts of the PI3-kinase inhibitor wortmannin. The compound had virtually no effect on tumor necrosis factor- and interleukin-1-induced NF-kappaB activities. Wortmannin did not inhibit tyrosine phosphorylation of IkappaB-alpha or alter the stability of the PI3-kinase complex but inhibited Akt kinase activation in response to pervanadate. Our data suggest that both the regulatory and the catalytic subunit of PI3-kinase play a role in NF-kappaB activation by the tyrosine phosphorylation-dependent pathway.
Figures
References
-
- Baeuerle P A, Baltimore D. Cell. 1996;87:13–20. - PubMed
-
- Baldwin A S., Jr Annu Rev Immunol. 1996;14:649–683. - PubMed
-
- Verma I M, Stevenson J K, Schwarz E M, Van Antwerp D, Miyamoto S. Genes Dev. 1995;9:2723–2735. - PubMed
-
- Bours V, Villalobos J, Burd P R, Kelly K, Siebenlist U. Nature (London) 1990;348:76–80. - PubMed
-
- Ghosh S, Gifford A M, Riviere L R, Tempst P, Nolan G P, Baltimore D. Cell. 1990;62:1019–1029. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
