

Moe1, a conserved protein in Schizosaccharomyces pombe, interacts with a Ras effector, Scd1, to affect proper spindle formation
- PMID: 9892665
- PMCID: PMC15168
- DOI: 10.1073/pnas.96.2.517
Moe1, a conserved protein in Schizosaccharomyces pombe, interacts with a Ras effector, Scd1, to affect proper spindle formation
Abstract
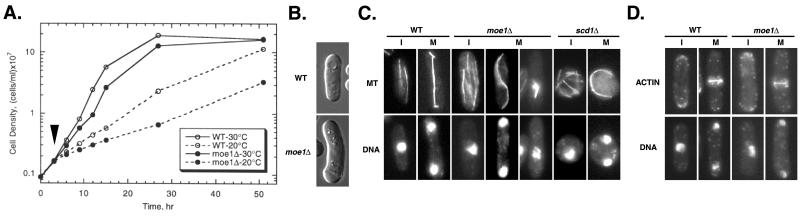
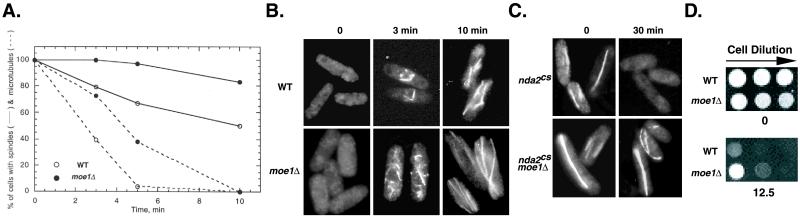
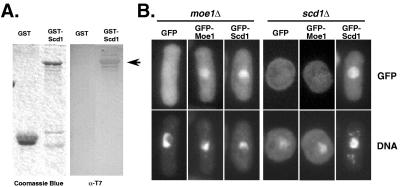
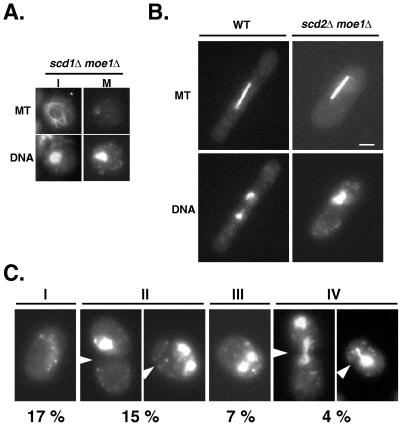
In fission yeast, Scd1/Ral1 is a putative guanine nucleotide exchange factor for Cdc42sp and also acts as a Ras1 effector necessary for the regulation of cytoskeleton organization. In this study, we have characterized a protein, Moe1, that binds directly to Scd1. A moe1 null (Delta) mutant exhibits numerous phenotypes indicative of abnormal microtubule functioning, including an abnormality in the spindle. moe1Delta mutants are resistant to microtubule destabilizing agents; moreover, moe1Delta rescued the growth defects of tubulin mutants containing unstable microtubules. These results suggest that Moe1 induces instability in microtubules. Biochemical and subcellular localization studies suggest that Moe1 and Scd1 colocalize in the nucleus. Furthermore, loss of function in Scd1 or Ras1 also induced abnormality in the spindle and is synthetically lethal with moe1Delta producing cells that lack a detectable spindle. These data demonstrate that Moe1 is a component of the Ras1 pathway necessary for proper spindle formation in the nucleus. Human and nematode Moe1 both can substitute for yeast Moe1, indicating that the function of Moe1 in spindle formation has been conserved substantially during evolution.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
