

Characterization of otoconin-95, the major protein of murine otoconia, provides insights into the formation of these inner ear biominerals
- PMID: 9892667
- PMCID: PMC15170
- DOI: 10.1073/pnas.96.2.529
Characterization of otoconin-95, the major protein of murine otoconia, provides insights into the formation of these inner ear biominerals
Abstract
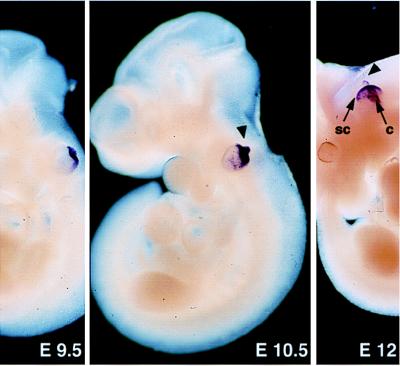
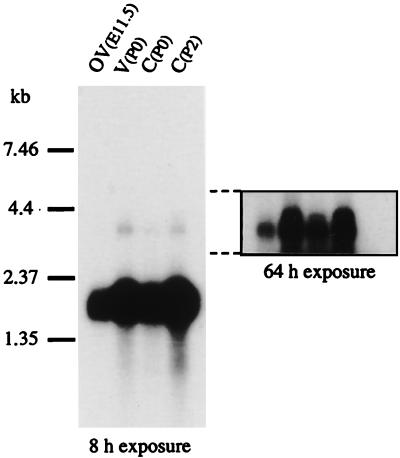
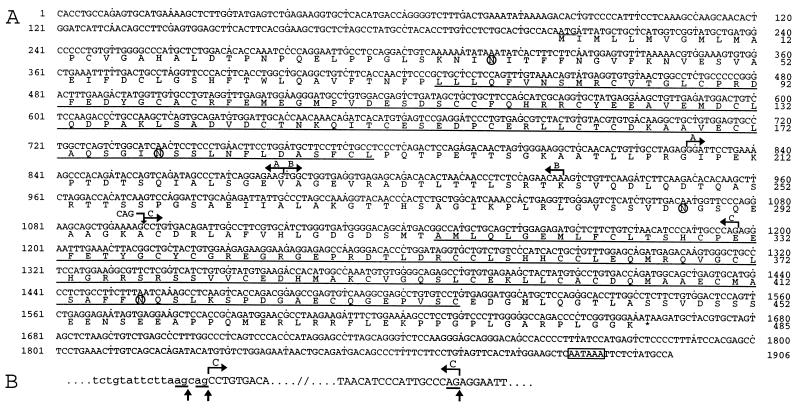
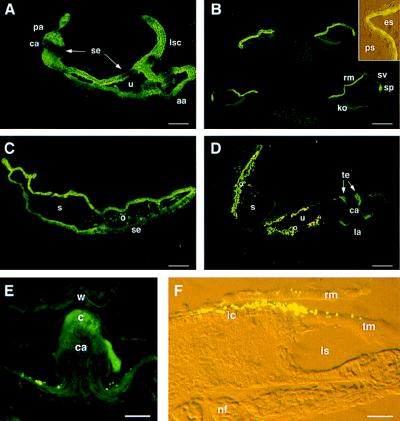
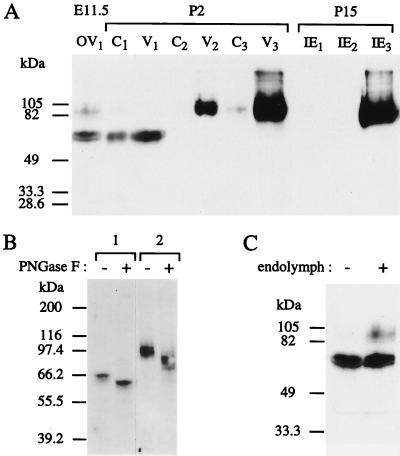
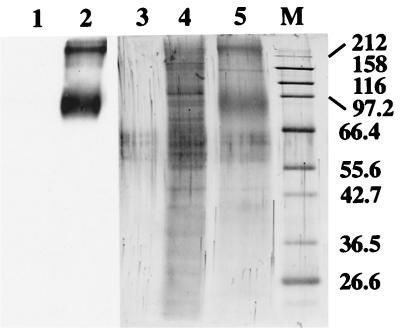
During the course of a study aimed at identifying inner ear-specific transcripts, a 1,906-bp murine cDNA predicted to encode a secreted 469-aa protein with two domains of homology with the secreted phospholipases A2 was isolated. This transcript is specifically expressed in the inner ear from embryonic day 9.5. The encoded 95-kDa glycoprotein is the major protein of the utricular and saccular otoconia and thus was named otoconin-95. By immunohistofluorescence, otoconin-95 also was detected in the cupulae of the semicircular canals and in previously undescribed transient granular structures of the cochlea. Otoconin-95 was found to be synthesized by various nonsensory cell types, but not by the supporting cells of the sensory epithelia, which produce the otoconial precursor vesicles. In addition, multiple isoforms generated by differential splicing were observed in different combinations during development. Based on the present results, we propose a model for the formation of the otoconia.
Figures
References
-
- Carlström D. Biol Bull. 1963;125:441–463.
-
- Marmo F, Balsamo G, Franco E. Cell Tissue Res. 1983;233:35–43. - PubMed
-
- Pote K G, Ross M D. Comp Biochem Physiol. 1991;98:287–295. - PubMed
-
- Pote K G, Ross M D. J Ultrastruct Mol Struct Res. 1986;95:61–70. - PubMed
-
- Pote K G, Hauer III C R, Michel H, Shabanowitz J, Hunt D F, Kretsinger R H. Biochemistry. 1993;32:5017–5024. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
