

The evolutionary origin of the protein-translocating channel of chloroplastic envelope membranes: identification of a cyanobacterial homolog
- PMID: 9892711
- PMCID: PMC15214
- DOI: 10.1073/pnas.96.2.784
The evolutionary origin of the protein-translocating channel of chloroplastic envelope membranes: identification of a cyanobacterial homolog
Abstract
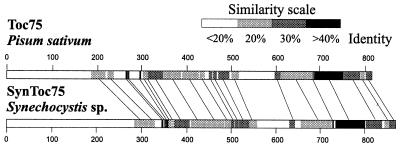
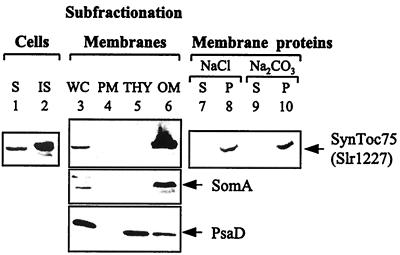
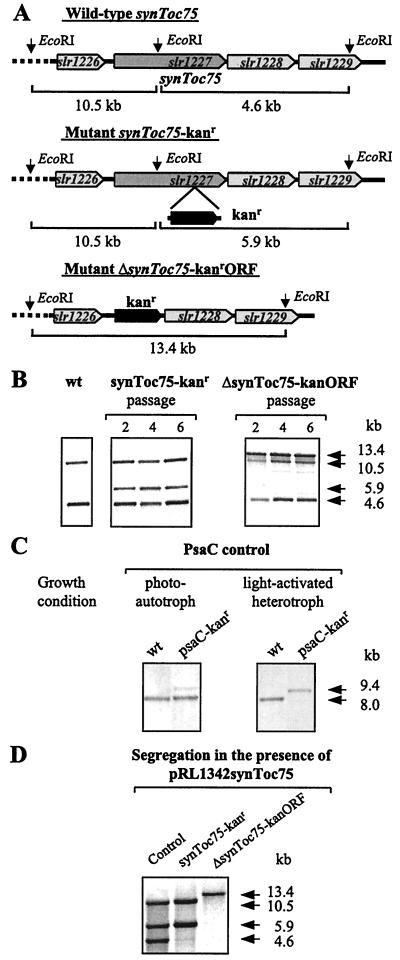
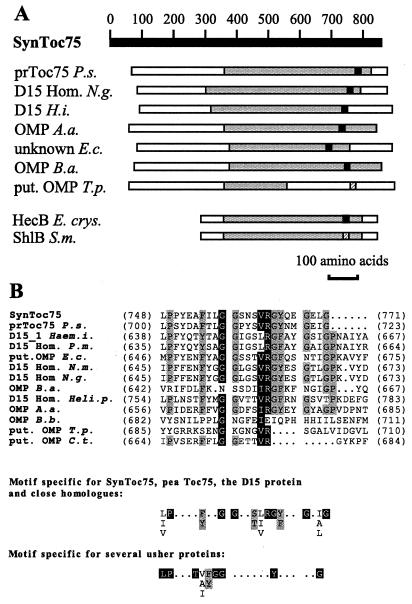
The known envelope membrane proteins of the chloroplastic protein import apparatus lack sequence similarity to proteins of other eukaryotic or prokaryotic protein transport systems. However, we detected a putative homolog of the gene encoding Toc75, the protein-translocating channel from the outer envelope membrane of pea chloroplasts, in the genome of the cyanobacterium Synechocystis sp. PCC 6803. We investigated whether the low sequence identity of 21% reflects a structural and functional relationship between the two proteins. We provide evidence that the cyanobacterial protein is also localized in the outer membrane. From this information and the similarity of the predicted secondary structures, we conclude that Toc75 and the cyanobacterial protein, referred to as SynToc75, are structural homologs. synToc75 is essential, as homozygous null mutants were not recovered after directed mutagenesis. Sequence analysis indicates that SynToc75 belongs to a family of outer membrane proteins from Gram-negative bacteria whose function is not yet known. However, we demonstrate that these proteins are related to a specific group of prokaryotic secretion channels that transfer virulence factors, such as hemolysins and adhesins, across the outer membrane.
Figures
Similar articles
-
Origin of a chloroplast protein importer.Proc Natl Acad Sci U S A. 1998 Dec 22;95(26):15831-6. doi: 10.1073/pnas.95.26.15831. Proc Natl Acad Sci U S A. 1998. PMID: 9861056 Free PMC article.
-
A Toc75-like protein import channel is abundant in chloroplasts.EMBO Rep. 2002 Jun;3(6):557-62. doi: 10.1093/embo-reports/kvf110. Epub 2002 May 24. EMBO Rep. 2002. PMID: 12034753 Free PMC article.
-
The chloroplastic protein translocation channel Toc75 and its paralog OEP80 represent two distinct protein families and are targeted to the chloroplastic outer envelope by different mechanisms.Plant J. 2004 Aug;39(3):354-65. doi: 10.1111/j.1365-313X.2004.02135.x. Plant J. 2004. PMID: 15255865
-
Arabidopsis genes encoding components of the chloroplastic protein import apparatus.Plant Physiol. 2001 Apr;125(4):1567-76. doi: 10.1104/pp.125.4.1567. Plant Physiol. 2001. PMID: 11299338 Free PMC article. Review.
-
Two evolutionarily conserved essential beta-barrel proteins in the chloroplast outer envelope membrane.Biosci Trends. 2009 Oct;3(5):168-78. Biosci Trends. 2009. PMID: 20103843 Review.
Cited by
-
On the origin of chloroplasts, import mechanisms of chloroplast-targeted proteins, and loss of photosynthetic ability - review.Folia Microbiol (Praha). 2009;54(4):303-21. doi: 10.1007/s12223-009-0048-z. Epub 2009 Oct 14. Folia Microbiol (Praha). 2009. PMID: 19826918 Review.
-
Leaf-specific upregulation of chloroplast translocon genes by a CCT motif-containing protein, CIA 2.Plant Cell. 2001 Sep;13(9):2053-61. doi: 10.1105/tpc.010148. Plant Cell. 2001. PMID: 11549763 Free PMC article.
-
A molecular-genetic study of the Arabidopsis Toc75 gene family.Plant Physiol. 2005 Jun;138(2):715-33. doi: 10.1104/pp.105.063289. Epub 2005 May 20. Plant Physiol. 2005. PMID: 15908591 Free PMC article.
-
Plastid origin and evolution: new models provide insights into old problems.Plant Physiol. 2011 Apr;155(4):1552-60. doi: 10.1104/pp.111.173500. Epub 2011 Feb 22. Plant Physiol. 2011. PMID: 21343425 Free PMC article. Review. No abstract available.
-
Borrelia burgdorferi locus BB0795 encodes a BamA orthologue required for growth and efficient localization of outer membrane proteins.Mol Microbiol. 2010 Feb;75(3):692-709. doi: 10.1111/j.1365-2958.2009.07015.x. Epub 2009 Dec 16. Mol Microbiol. 2010. PMID: 20025662 Free PMC article.
References
-
- Schimper A F W. Bot Z. 1883;41:105–114.
-
- Mereschowsky C. Biol Zentralbl. 1905;25:593–604. , 689–691.
-
- Gray M W. Int Rev Cytol. 1992;141:233–357. - PubMed
-
- Martin W, Stoebe B, Goremykin V, Hansmann S, Hasegawa M, Kowallik K V. Nature (London) 1998;393:162–165. - PubMed
-
- Keegstra K. Cell. 1989;56:247–253. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
