

The effect of antibody-dependent enhancement on the transmission dynamics and persistence of multiple-strain pathogens
- PMID: 9892712
- PMCID: PMC15215
- DOI: 10.1073/pnas.96.2.790
The effect of antibody-dependent enhancement on the transmission dynamics and persistence of multiple-strain pathogens
Abstract
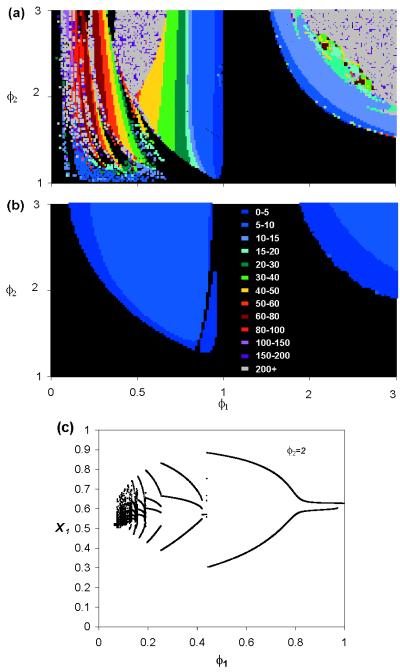
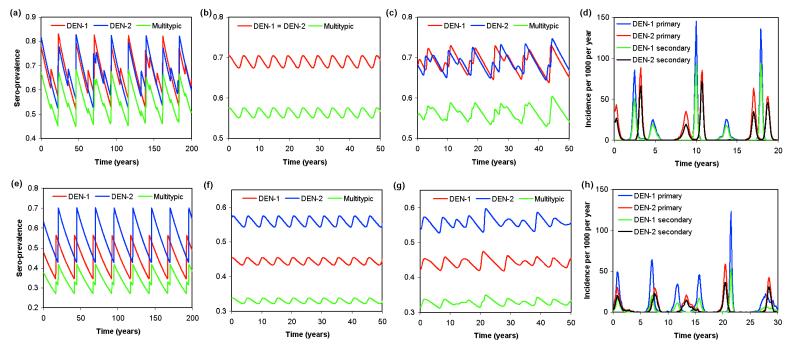
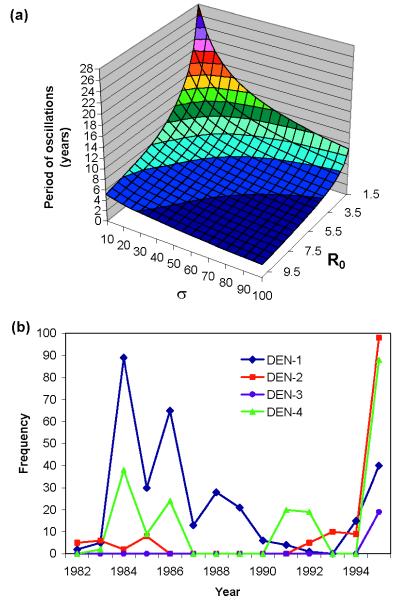
Cross-reactive antibodies produced by a mammalian host during infection by a particular microparasitic strain usually have the effect of reducing the probability of the host being infected by a different, but closely related, pathogen strain. Such cross-reactive immunological responses thereby induce between-strain competition within the pathogen population. However, in some cases such as dengue virus, evidence suggests that cross-reactive antibodies act to enhance rather than restrict the severity of a subsequent infection by another strain. This cooperative mechanism is thought to explain why pre-existing immunity to dengue virus is an important risk factor for the development of severe disease (i.e., dengue shock syndrome and dengue hemorrhagic fever). In this paper, we explore the effect of antibody-dependent enhancement on the transmission dynamics of multistrain pathogen populations. We show that enhancement frequently may generate complex and persistent cyclical or chaotic epidemic behavior. Furthermore, enhancement acts to permit the coexistence of all strains where in its absence only one or a subset would persist.
Figures
References
-
- Adler F R, Brunet R C. Theor Popul Biol. 1991;40:369–410. - PubMed
-
- Gupta S, Swinton J, Anderson R M. Proc R Soc London Ser B. 1994;256:231–238. - PubMed
-
- Gupta S, Maiden M C J, Feavers I M, Nee S, May R M, Anderson R M. Nat Med. 1996;2:437–442. - PubMed
-
- Andreasen V, Lin J, Levin S A. J Math Biol. 1997;35:825–842. - PubMed
-
- Gupta S, Ferguson N M, Anderson R M. Science. 1998;280:912–915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
