

Mutant huntingtin expression in clonal striatal cells: dissociation of inclusion formation and neuronal survival by caspase inhibition
- PMID: 9920660
- PMCID: PMC6782141
- DOI: 10.1523/JNEUROSCI.19-03-00964.1999
Mutant huntingtin expression in clonal striatal cells: dissociation of inclusion formation and neuronal survival by caspase inhibition
Abstract
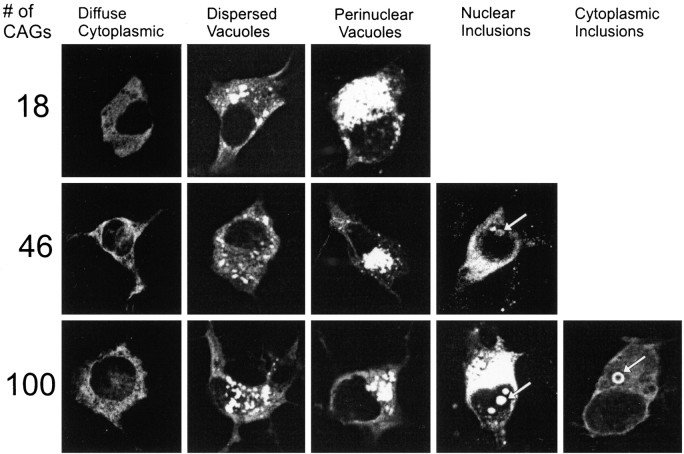
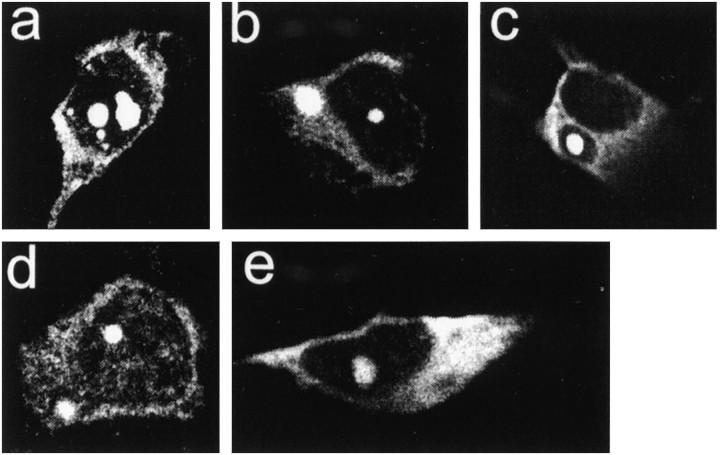
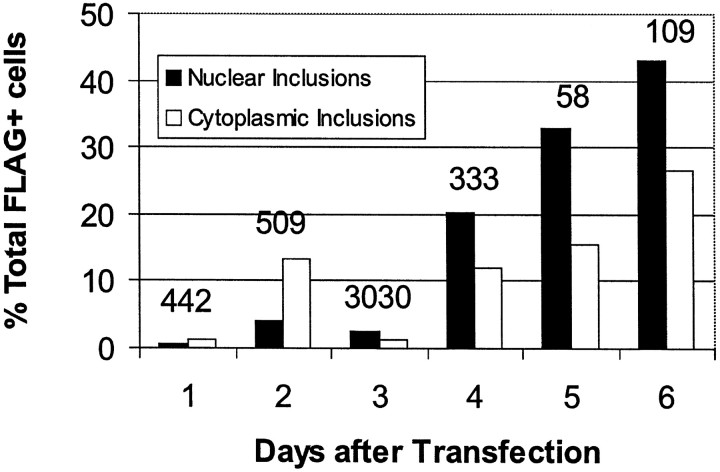
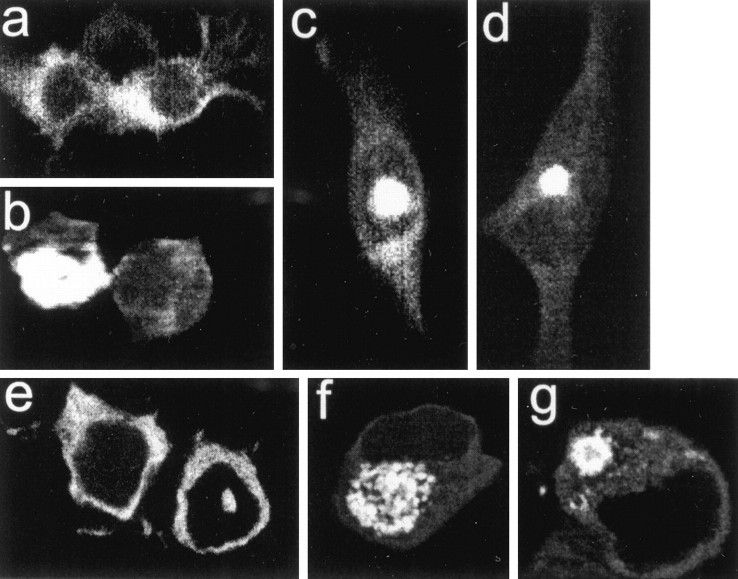
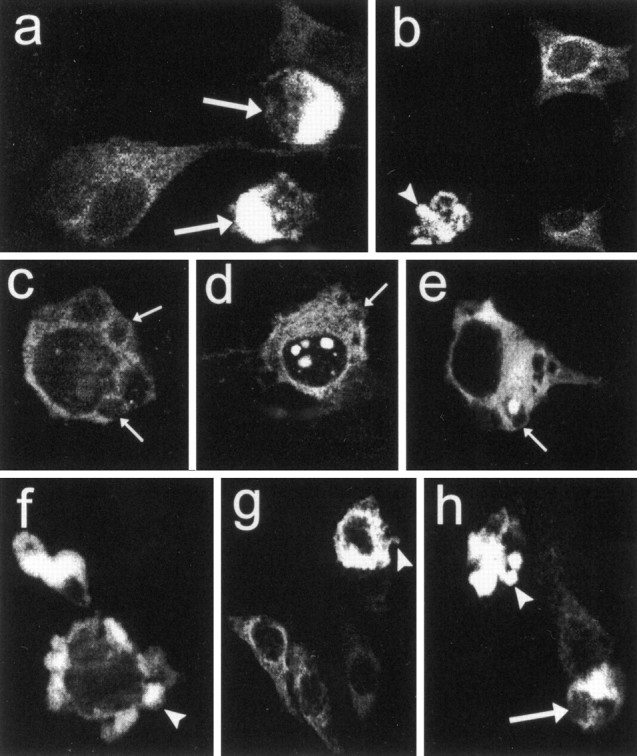
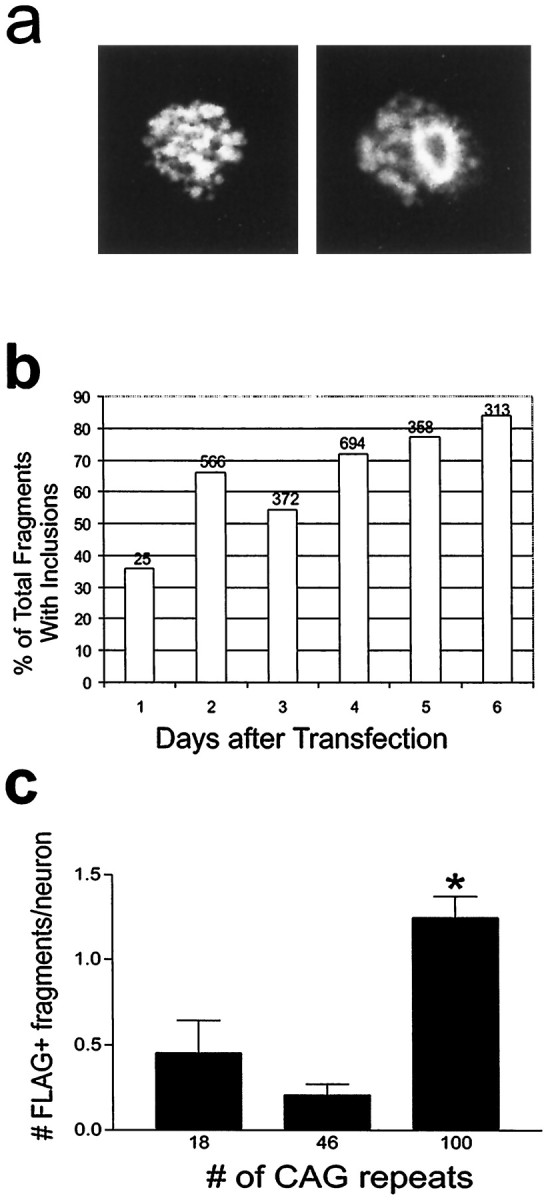
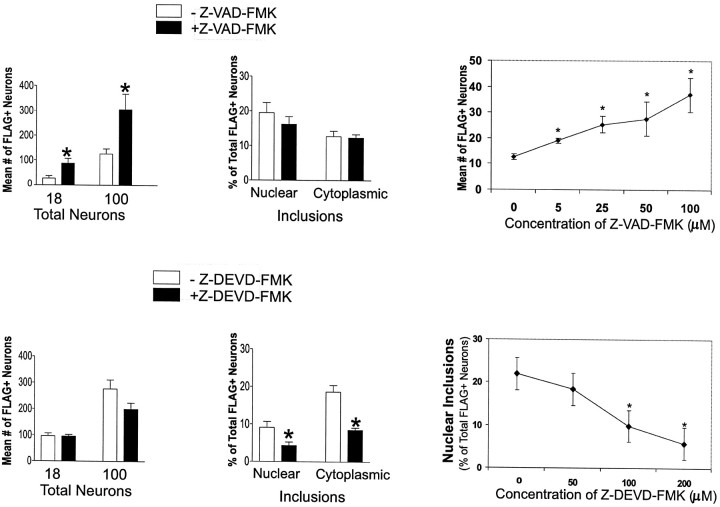
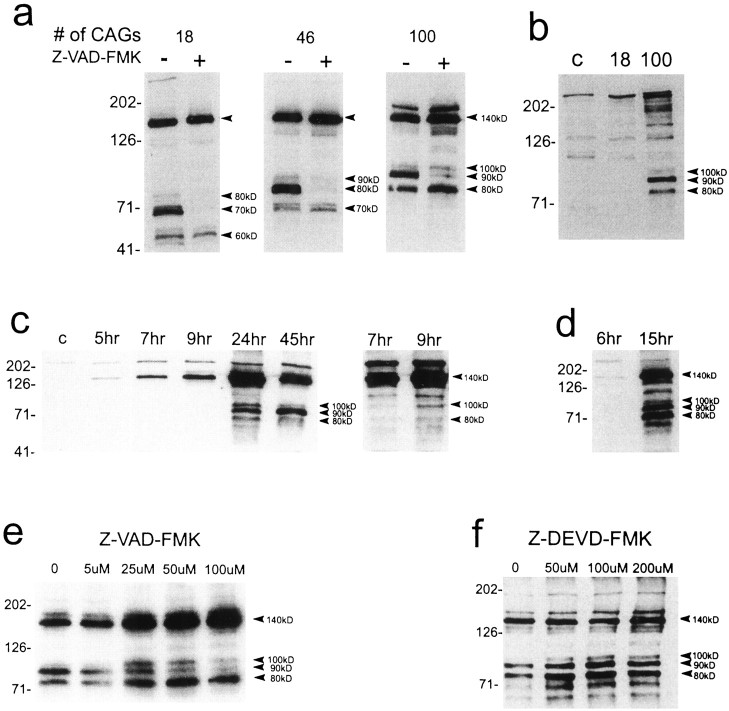
Neuronal intranuclear inclusions are found in the brains of patients with Huntington's disease and form from the polyglutamine-expanded N-terminal region of mutant huntingtin. To explore the properties of inclusions and their involvement in cell death, mouse clonal striatal cells were transiently transfected with truncated and full-length human wild-type and mutant huntingtin cDNAs. Both normal and mutant proteins localized in the cytoplasm, and infrequently, in dispersed and perinuclear vacuoles. Only mutant huntingtin formed nuclear and cytoplasmic inclusions, which increased with polyglutamine expansion and with time after transfection. Nuclear inclusions contained primarily cleaved N-terminal products, whereas cytoplasmic inclusions contained cleaved and larger intact proteins. Cells with wild-type or mutant protein had distinct apoptotic features (membrane blebbing, shrinkage, cellular fragmentation), but those with mutant huntingtin generated the most cell fragments (apoptotic bodies). The caspase inhibitor Z-VAD-FMK significantly increased cell survival but did not diminish nuclear and cytoplasmic inclusions. In contrast, Z-DEVD-FMK significantly reduced nuclear and cytoplasmic inclusions but did not increase survival. A series of N-terminal products was formed from truncated normal and mutant proteins and from full-length mutant huntingtin but not from full-length wild-type huntingtin. One prominent N-terminal product was blocked by Z-VAD-FMK. In summary, the formation of inclusions in clonal striatal cells corresponds to that seen in the HD brain and is separable from events that regulate cell death. N-terminal cleavage may be linked to mutant huntingtin's role in cell death.
Figures
References
-
- Becher MW, Kotzuk JA, Sharp AH, Davies SW, Bates GP, Price DL, Ross CA. Intranuclear neuronal inclusions in Huntington’s disease and dentatorubral and pallidoluysian atrophy: correlation between the density of inclusions and IT15 CAG triplet repeat length. Neurobiol Dis. 1998;4:387–397. - PubMed
-
- Choi HK, Won LA, Kontur PJ, Hammond DN, Fox AP, Wainer B, Hoffman PC, Heller A. Immortalization of embryonic mesencephalic dopaminergic neurons by somatic cell fusion. Brain Res. 1991;552:67–76. - PubMed
-
- Cooper AJ, Sheu KF, Burke JR, Onodera O, Strittmatter WJ, Roses AD, Blass JP. Polyglutamine domains are substrates of tissue transglutaminase: does transglutaminase play a role in expanded CAG/poly-Q neurodegenerative diseases? J Neurochem. 1997;69:431–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources