

Biophysical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): molecular size, glycosylation pattern, and engineering of proteolytic resistance
- PMID: 9925554
- PMCID: PMC91033
- DOI: 10.1128/AEM.65.2.359-366.1999
Biophysical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): molecular size, glycosylation pattern, and engineering of proteolytic resistance
Abstract
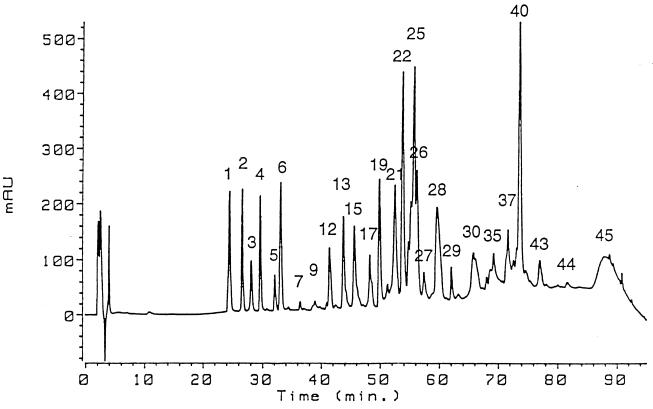
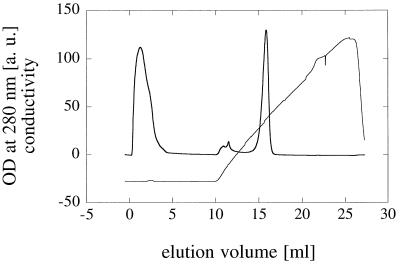
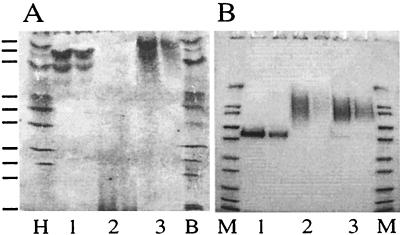
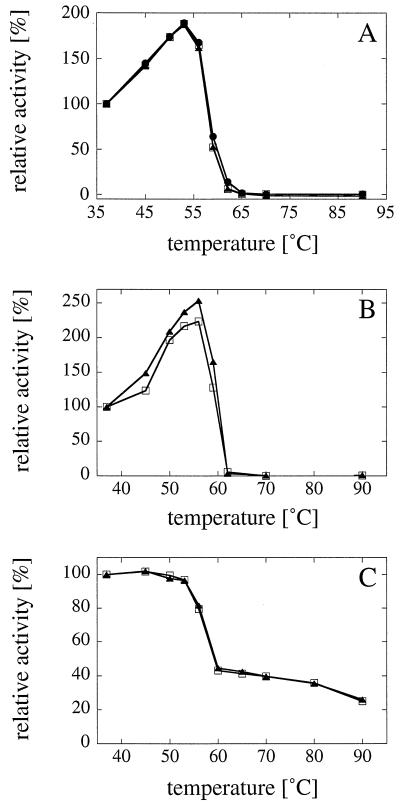
Phytases (myo-inositol hexakisphosphate phosphohydrolases) are found naturally in plants and microorganisms, particularly fungi. Interest in these enzymes has been stimulated by the fact that phytase supplements increase the availability of phosphorus in pig and poultry feed and thereby reduce environmental pollution due to excess phosphate excretion in areas where there is intensive livestock production. The wild-type phytases from six different fungi, Aspergillus niger, Aspergillus terreus, Aspergillus fumigatus, Emericella nidulans, Myceliophthora thermophila, and Talaromyces thermophilus, were overexpressed in either filamentous fungi or yeasts and purified, and their biophysical properties were compared with those of a phytase from Escherichia coli. All of the phytases examined are monomeric proteins. While E. coli phytase is a nonglycosylated enzyme, the glycosylation patterns of the fungal phytases proved to be highly variable, differing for individual phytases, for a given phytase produced in different expression systems, and for individual batches of a given phytase produced in a particular expression system. Whereas the extents of glycosylation were moderate when the fungal phytases were expressed in filamentous fungi, they were excessive when the phytases were expressed in yeasts. However, the different extents of glycosylation had no effect on the specific activity, the thermostability, or the refolding properties of individual phytases. When expressed in A. niger, several fungal phytases were susceptible to limited proteolysis by proteases present in the culture supernatant. N-terminal sequencing of the fragments revealed that cleavage invariably occurred at exposed loops on the surface of the molecule. Site-directed mutagenesis of A. fumigatus and E. nidulans phytases at the cleavage sites yielded mutants that were considerably more resistant to proteolytic attack. Therefore, engineering of exposed surface loops may be a strategy for improving phytase stability during feed processing and in the digestive tract.
Figures
References
-
- Davis R W, Botstein D, Roth J R. A manual for genetic engineering: advanced bacterial genetics. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1980. pp. 120–123.
-
- Gellissen G, Janowicz Z A, Merckelbach A, Piontek M, Keup P, Weydemann U, Hollenberg C P, Strasser A W M. Heterologous gene expression in Hansenula polymorpha: efficient secretion of glucoamylase. Bio/Technology. 1991;9:291–295. - PubMed
-
- Gibson D M, Ullah A H J. Purification and characterization of phytase from cotyledons of germinating soybean seeds. Arch Biochem Biophys. 1988;260:503–513. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
