

Generation of conditional mutants in higher eukaryotes by switching between the expression of two genes
- PMID: 9927685
- PMCID: PMC15342
- DOI: 10.1073/pnas.96.3.1013
Generation of conditional mutants in higher eukaryotes by switching between the expression of two genes
Abstract
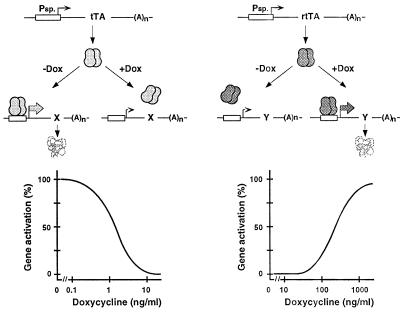
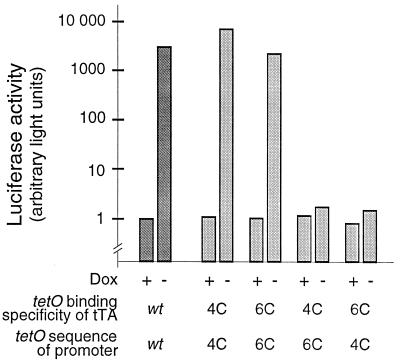
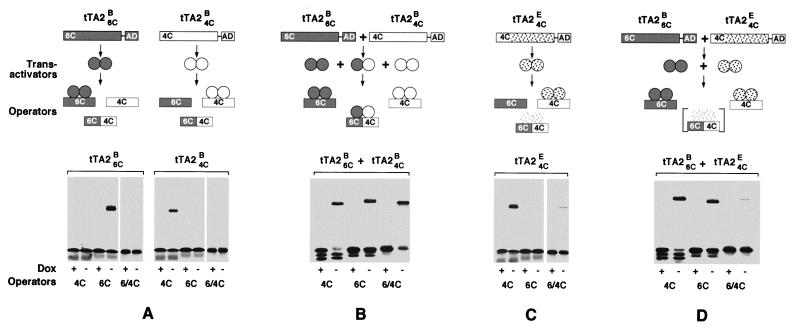
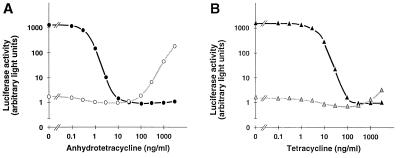
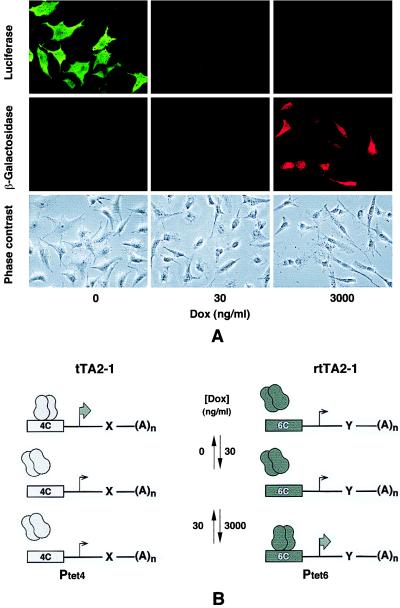
A regulatory system for the in-depth study of gene functions in higher eukaryotic cells has been developed. It is based on the tetracycline-controlled transactivators and reverse tTA, which were remodeled to discriminate efficiently between two different promoters. The system permits one to control reversibly the activity of two genes, or two alleles of a gene, in a mutually exclusive way, and also allows one to abrogate the activities of both. This dual regulatory circuit, which can be operated by a single effector substance such as doxycycline, overcomes limitations of conventional genetic approaches. The conditional mutants that can now be generated will be useful for the study of gene function in vitro and in vivo. In addition, the system may be of value for a variety of practical applications, including gene therapy.
Figures
Comment in
-
Tet B or not tet B: advances in tetracycline-inducible gene expression.Proc Natl Acad Sci U S A. 1999 Feb 2;96(3):797-9. doi: 10.1073/pnas.96.3.797. Proc Natl Acad Sci U S A. 1999. PMID: 9927646 Free PMC article. Review. No abstract available.
References
-
- Gossen M, Freundlieb S, Bender G, Müller G, Hillen W, Bujard H. Science. 1995;268:1766–1769. - PubMed
-
- Freundlieb S, Baron U, Bonin A, Gossen M, Bujard H. Methods Enzymol. 1997;283:159–172. - PubMed
-
- Camacho-Vanegas O, Mannucci L, Amaldi F. In Vitro Cell Dev Biol Anim. 1998;34:14–15. - PubMed
-
- Garí E, Piedrafita L, Aldea M, Herrero E. Yeast. 1997;13:837–848. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
