

Regulation of CR3 (CD11b/CD18)-dependent natural killer (NK) cell cytotoxicity by tumour target cell MHC class I molecules
- PMID: 9933447
- PMCID: PMC1905157
- DOI: 10.1046/j.1365-2249.1999.00800.x
Regulation of CR3 (CD11b/CD18)-dependent natural killer (NK) cell cytotoxicity by tumour target cell MHC class I molecules
Abstract
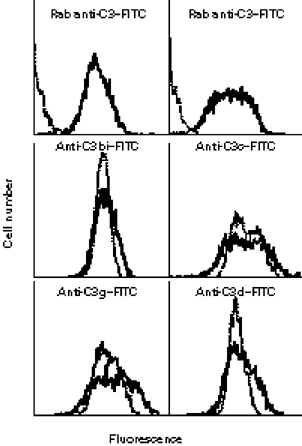
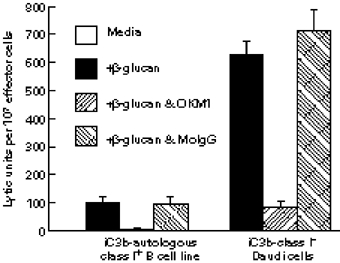
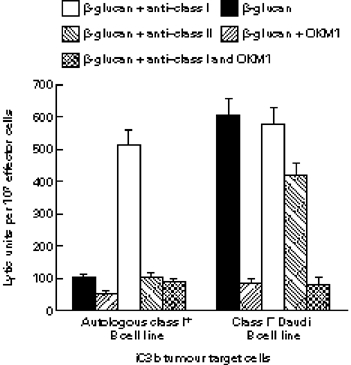
Phagocyte and NK cell CR3 functions as both an adhesion molecule and an iC3b receptor mediating cytotoxic responses to microorganisms. Cytotoxic activation of iC3b receptor function requires ligation of both a CD11b I-domain site for iC3b and a lectin site located in the C-terminus of CD11b. Because tumours lack the CR3-binding polysaccharides of bacteria and fungi, iC3b-opsonized tumours do not stimulate CR3-dependent cytotoxicity. Previous studies showed that NK cells could be induced to kill iC3b-opsonized tumours with small soluble beta-glucans that bound with high affinity to CR3, bypassing the absence of similar polysaccharides on tumour membranes. Because CR3 signalling requires several tyrosine phosphorylation events, it appeared possible that CR3-dependent killing of autologous tumour cells might be suppressed by NK cell inhibitory receptors for MHC class I (KIR and CD94/NKG2) whose action involves recruitment of SHP-1 and SHP-2 tyrosine phosphatases. In the current study, Epstein-Barr virus (EBV)-transformed B cells were used as targets following opsonization with iC3b. Soluble beta-glucan primed CR3 for killing of iC3b-coated B cells, but autologous class I-bearing targets were 84% more resistant than class I-deficient Daudi cells. Blockade of target cell class I with a MoAb specific for a domain recognized by both KIR and CD94/NKG2 resulted in comparable killing of class I+ B cells. By contrast, another MoAb to class II had no effect on cytotoxicity. These data suggest that NK cell recognition of class I suppresses CR3/tyrosine kinase-dependent cytotoxicity in the same way as it suppresses cytotoxicity mediated by other tyrosine kinase-linked receptors such as FcgammaRIIIA (CD16).
Figures
References
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76:301–14. - PubMed
-
- Hogg N, Berlin C. Structure and function of adhesion receptors in leukocyte trafficking. Immunol Today. 1995;16:327–30. - PubMed
-
- Sugimori T, Griffith DL, Arnaout MA. Emerging paradigms of integrin ligand binding and activation. Kidney Int. 1997;51:1454–62. - PubMed
-
- Petty HR, Todd RF. Receptor–receptor interactions of complement receptor type 3 in neutrophil membranes. J Leuk Biol. 1993;54:492–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
