

Complementary DNA cloning and characterization of ferredoxin localized in bundle-sheath cells of maize leaves
- PMID: 9952443
- PMCID: PMC32124
- DOI: 10.1104/pp.119.2.481
Complementary DNA cloning and characterization of ferredoxin localized in bundle-sheath cells of maize leaves
Abstract
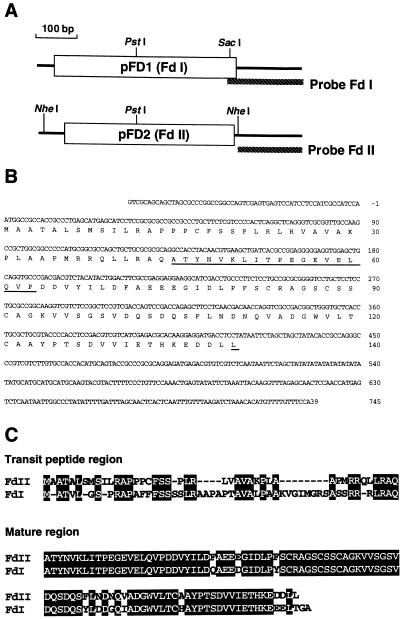
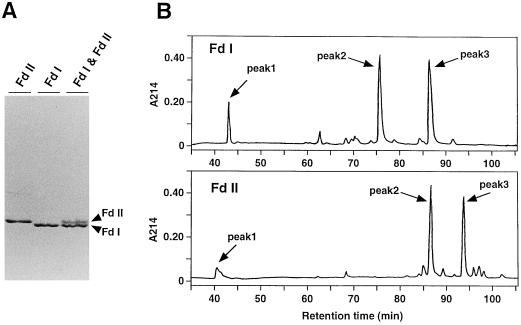
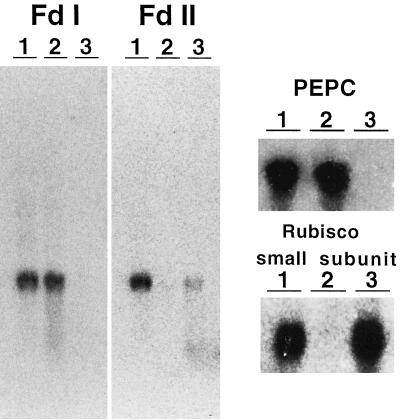
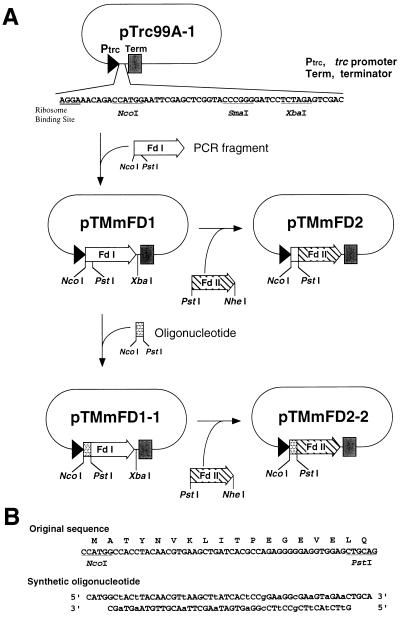
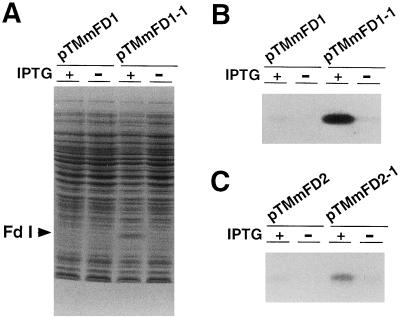
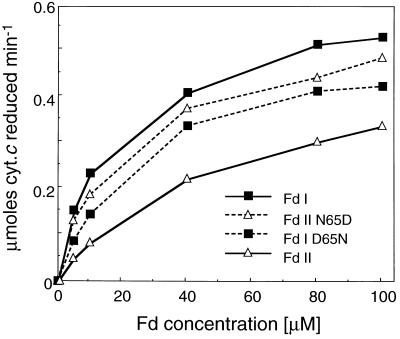
In maize (Zea mays L.) two leaf-specific ferredoxin (Fd) isoproteins, Fd I and Fd II, are distributed differentially in mesophyll and bundle-sheath cells. A novel cDNA encoding the precursor of Fd II (pFD2) was isolated by heterologous hybridization using a cDNA for Fd I (pFD1) as a probe. The assignment of the cDNAs to the Fds was verified by capillary liquid-chromatography/electrospray ionization-mass spectrometry. RNA-blot analysis demonstrated that transcripts for Fd I and Fd II accumulated specifically in mesophyll and bundle-sheath cells, respectively. The mature regions of pFD1 and pFD2 were expressed in Escherichia coli as functional Fds. Fd I and Fd II had similar redox potentials of -423 and -406 mV, respectively, but the Km value of Fd-NADP+ reductase for Fd II was about 3-fold larger than that for Fd I. Asparagine at position 65 of Fd II is a unique residue compared with Fd I and other Fds from various plants, which have aspartic acid or glutamic acid at the corresponding position as an electrostatic interaction site with Fd-NADP+ reductase. Substitution of asparagine-65 with aspartic acid increased the affinity of Fd II with Fd-NADP+ reductase to a level comparable to that of Fd I. These structural and functional differences of Fd I and Fd II may be related to their cell-specific expression in the leaves of a C4 plant.
Figures
Similar articles
-
Differential interaction of maize root ferredoxin:NADP(+) oxidoreductase with photosynthetic and non-photosynthetic ferredoxin isoproteins.Plant Physiol. 2000 Jul;123(3):1037-45. doi: 10.1104/pp.123.3.1037. Plant Physiol. 2000. PMID: 10889253 Free PMC article.
-
A post genomic characterization of Arabidopsis ferredoxins.Plant Physiol. 2004 Jan;134(1):255-64. doi: 10.1104/pp.103.032755. Epub 2003 Dec 18. Plant Physiol. 2004. PMID: 14684843 Free PMC article.
-
Differential response of genes for ferredoxin and ferredoxin:NADP+ oxidoreductase to nitrate and light in maize leaves.J Plant Physiol. 2003 Jan;160(1):65-70. doi: 10.1078/0176-1617-00919. J Plant Physiol. 2003. PMID: 12685047
-
Amino acid residues responsible for the different pH dependency of cell-specific ferredoxins in the electron transfer reaction with ferredoxin-NADP+ reductase from maize leaves.J Biochem. 2024 Sep 3;176(3):237-244. doi: 10.1093/jb/mvae043. J Biochem. 2024. PMID: 38861409
-
The end of the line: can ferredoxin and ferredoxin NADP(H) oxidoreductase determine the fate of photosynthetic electrons?Curr Protein Pept Sci. 2014;15(4):385-93. doi: 10.2174/1389203715666140327113733. Curr Protein Pept Sci. 2014. PMID: 24678667 Free PMC article. Review.
Cited by
-
Differential interaction of maize root ferredoxin:NADP(+) oxidoreductase with photosynthetic and non-photosynthetic ferredoxin isoproteins.Plant Physiol. 2000 Jul;123(3):1037-45. doi: 10.1104/pp.123.3.1037. Plant Physiol. 2000. PMID: 10889253 Free PMC article.
-
The role of 5'-adenylylsulfate reductase in controlling sulfate reduction in plants.Photosynth Res. 2005 Dec;86(3):309-23. doi: 10.1007/s11120-005-9006-z. Epub 2005 Nov 15. Photosynth Res. 2005. PMID: 16328785 Review.
-
RNA-sequencing transcriptome analysis of Avicennia marina (Forsk.) Vierh. leaf epidermis defines tissue-specific transcriptional response to salinity treatment.Sci Rep. 2023 May 10;13(1):7614. doi: 10.1038/s41598-023-34095-x. Sci Rep. 2023. PMID: 37165000 Free PMC article.
-
FdC1, a novel ferredoxin protein capable of alternative electron partitioning, increases in conditions of acceptor limitation at photosystem I.J Biol Chem. 2011 Jan 7;286(1):50-9. doi: 10.1074/jbc.M110.161562. Epub 2010 Oct 21. J Biol Chem. 2011. PMID: 20966083 Free PMC article.
-
Light-Independent Nitrogen Assimilation in Plant Leaves: Nitrate Incorporation into Glutamine, Glutamate, Aspartate, and Asparagine Traced by 15N.Plants (Basel). 2020 Oct 2;9(10):1303. doi: 10.3390/plants9101303. Plants (Basel). 2020. PMID: 33023108 Free PMC article. Review.
References
-
- Arnon DI. The discovery of ferredoxin: the photosynthetic path. Trends Biochem Sci. 1989;13:30–33. - PubMed
-
- Asada K, Herber U, Schreiber U. Electron flow to the intersystem chain from stromal components and cyclic electron flow in maize chloroplasts, as detected in intact leaves by monitoring redox change of P700 and chlorophyll fluorescence. Plant Cell Physiol. 1993;34:39–50.
-
- Ausbel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (1991) Mutagenesis by the polymerase chain reaction. In Current Protocols in Molecular Biology. John Wiley & Sons, New York, pp 1–9
-
- Crestfield AM, Moore S, Stein WH. The preparation and enzymatic hydrolysis of reduced and S-carboxymethylated proteins. J Biol Chem. 1963;238:622–627. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
