

Ectopic expression of the maize homeobox gene liguleless3 alters cell fates in the leaf
- PMID: 9952462
- PMCID: PMC32143
- DOI: 10.1104/pp.119.2.651
Ectopic expression of the maize homeobox gene liguleless3 alters cell fates in the leaf
Abstract
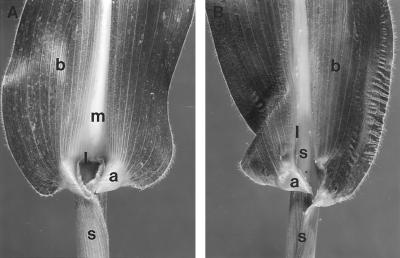
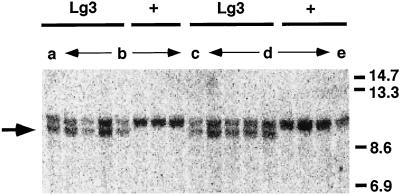
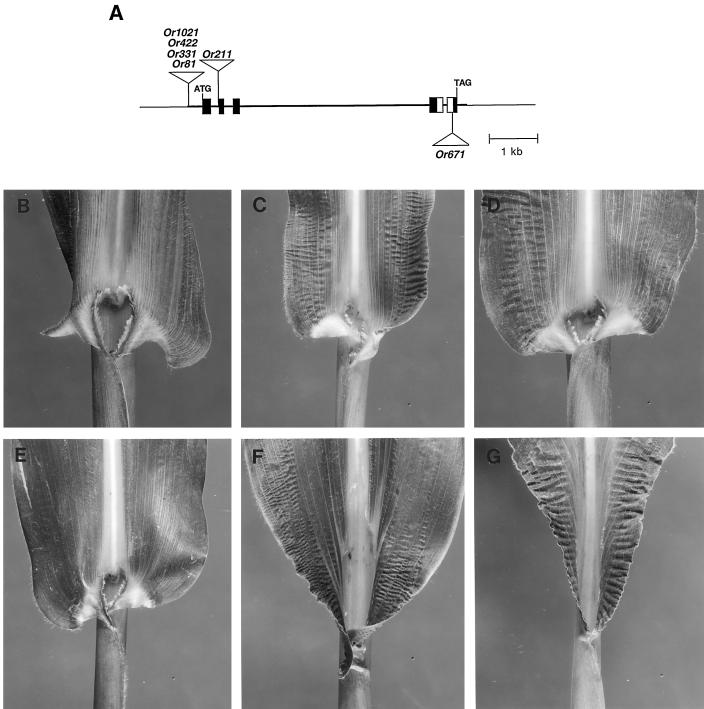
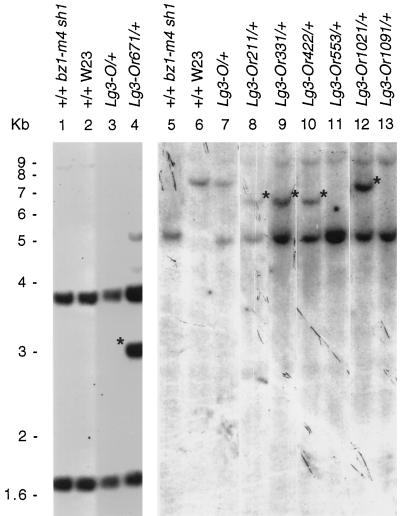
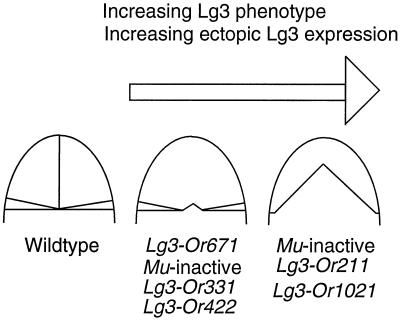
The semidominant mutation Liguleless3-O (Lg3-O) causes a blade-to-sheath transformation at the midrib region of the maize (Zea mays L.) leaf. We isolated a full-length lg3 cDNA containing a knotted1-like family homeobox. Six Lg3-O partial revertant alleles caused by insertion of a Mutator (Mu) transposon and two deletion derivatives were isolated and used to verify that our knotted1-like cDNA corresponds to the LG3 message. In wild-type plants the LG3 mRNA is expressed in apical regions but is not expressed in leaves. In mutant plants harboring any of three dominant lg3 alleles (Lg3-O, -Mlg, and -347), LG3 mRNA is expressed in leaf sheath tissue, indicating that the Lg3 phenotype is due to ectopic expression of the gene. The Lg3-O revertant alleles represent two classes of Lg3 phenotypes that correlate well with the level of ectopic Lg3 expression. High levels of ectopic LG3 mRNA expression results in a severe Lg3 phenotype, whereas weak ectopic Lg3 expression results in a mild Lg3 phenotype. We propose that ectopic Lg3 expression early in leaf development causes the blade-to-sheath transformation, but the level of expression determines the extent of the transformation.
Figures




Similar articles
-
Sectors expressing the homeobox gene liguleless3 implicate a time-dependent mechanism for cell fate acquisition along the proximal-distal axis of the maize leaf.Development. 1997 Dec;124(24):5097-106. doi: 10.1242/dev.124.24.5097. Development. 1997. PMID: 9362467
-
Regulation and a conserved intron sequence of liguleless3/4 knox class-I homeobox genes in grasses.Planta. 2004 Jun;219(2):359-68. doi: 10.1007/s00425-004-1233-6. Epub 2004 Mar 19. Planta. 2004. PMID: 15034715
-
Mosaic analysis of the liguleless3 mutant phenotype in maize by coordinate suppression of mutator-insertion alleles.Genetics. 1996 May;143(1):489-503. doi: 10.1093/genetics/143.1.489. Genetics. 1996. PMID: 8722798 Free PMC article.
-
Plant morphogenesis. More knots untied.Curr Biol. 1994 Jun 1;4(6):529-31. doi: 10.1016/s0960-9822(00)00115-9. Curr Biol. 1994. PMID: 7922374 Review.
-
Genetic and molecular analysis of patterning in plant development.ASGSB Bull. 1995 Oct;8(2):29-37. ASGSB Bull. 1995. PMID: 11538548 Review.
Cited by
-
A dynamic regulome of shoot-apical-meristem-related homeobox transcription factors modulates plant architecture in maize.Genome Biol. 2024 Sep 19;25(1):245. doi: 10.1186/s13059-024-03391-8. Genome Biol. 2024. PMID: 39300560 Free PMC article.
-
Genetic basis controlling rice plant architecture and its modification for breeding.Breed Sci. 2023 Mar;73(1):3-45. doi: 10.1270/jsbbs.22088. Epub 2023 Mar 29. Breed Sci. 2023. PMID: 37168811 Free PMC article.
-
Regulation of SHOOT MERISTEMLESS genes via an upstream-conserved noncoding sequence coordinates leaf development.Proc Natl Acad Sci U S A. 2007 Oct 2;104(40):15953-8. doi: 10.1073/pnas.0707577104. Epub 2007 Sep 26. Proc Natl Acad Sci U S A. 2007. PMID: 17898165 Free PMC article.
-
Developmental genetics of maize vegetative shoot architecture.Mol Breed. 2021 Feb 22;41(3):19. doi: 10.1007/s11032-021-01208-1. eCollection 2021 Mar. Mol Breed. 2021. PMID: 37309417 Free PMC article.
-
Pathogen-induced binding of the soybean zinc finger homeodomain proteins GmZF-HD1 and GmZF-HD2 to two repeats of ATTA homeodomain binding site in the calmodulin isoform 4 (GmCaM4) promoter.Nucleic Acids Res. 2007;35(11):3612-23. doi: 10.1093/nar/gkm273. Epub 2007 May 7. Nucleic Acids Res. 2007. PMID: 17485478 Free PMC article.
References
-
- Barton MK, Poethig S. Formation of the shoot apical meristem in Arabidopsis thaliana: an analysis of development in the wild type and shoot meristemless mutant. Development. 1993;119:823–831.
-
- Becraft PW, Bongard-Pierce DK, Sylvester AW, Poethig RS, Freeling M. The liguleless1 gene acts tissue specifically in maize leaf development. Dev Biol. 1990;141:220–232. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
